分子植物育种
(
网络版
), 2016
年
,
第
14
卷
,
第
1008
-
1015
页
Fenzi Zhiwu Yuzhong (Online), 2016, Vol.14, 1008
-
1015
Copyright © 2016 BioPublisher 1013
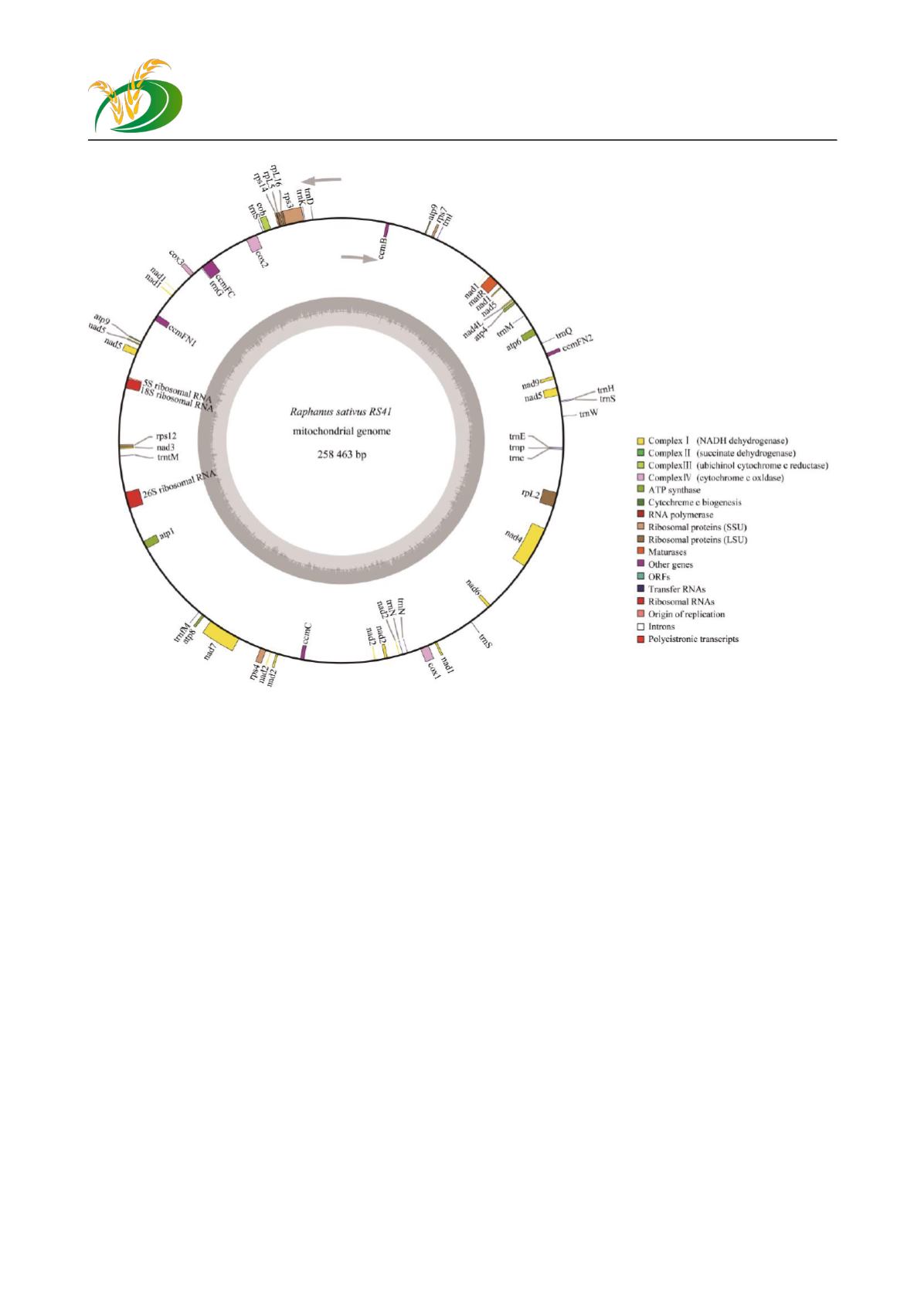
图
5
萝卜
RS41
的线粒体环形基因组
Figure5 Gene map for RS41’s mitochondrial
另一方面,由于本研究是基于有参考基因组的
组装。因此并未建立
mate-pair
文库、
fosmid
文库或
进行三代
Pacbio
测序。如果是全新的
De novo
组装
则必须有
mate-pair
文库、
fosmid
文库或三代测序的
数据支持才能进行有效组装。因此应针对不同的组
装方法采取不同的建库策略。
2.2
线粒体与叶绿体的共有序列
在线粒体与叶绿体的组装中,我们发现两者存
在
6
处大小不一的共有序列,这些共有序列存在较
高的相似性。此处的
Contigs
难以确定归属,影响
组装效果。特别是在线粒体组装,去除叶绿体
Reads
时应以本样品组装的叶绿体基因组为参考,进行严
谨比对
(mismatch=0)
,这样重复序列区段可以连接
到线粒体基因组上,否则影响线粒体的组装。
2.3
关于测序深度
从我们的组装情况看,对于像叶绿体和线粒体
这样大小的基因组,能够达到
800
倍左右的测序深
度,就比较好组装了,本研究的叶绿体组装就比较
顺利,如果没有重复序列,组装的
Contigs
将会长
而且数目少。而对于线粒体来说,测序深度仅
110
倍左右,组装难度大,组装出的
Contigs
多,存在
较多
Gap
,如果没有比较相近的参照基因组,组装
工作会非常困难。
2.4
基因注释
组装完毕后我们对各基因组均进行了基因注
释,基因注释的结果呈现出叶绿体、线粒体作为自
主或半自主细胞器的特征
(Ris and Plaut, 1962;
Saccone et al., 2000)
。而基因组之间的差异包括
SNP
及结构变异是否分布于
CDS
区域需要进一步的生
物信息学分析来加以阐述。
3
材料与方法
3.1
实验材料
萝卜胞质雄性不育材料
RS41
、
RS42
、
RS43
,
未知不育类型
RS44
及可育细胞质材料
RS45
由本
课题组收集。试验材料种植于课题组试验田,田间
管理按常规方法进行,待植株发育至
10
片叶时,


