Basic HTML Version


周坤华等
, 2011,
辣椒属栽培种、野生种和种间杂交后代的
SRAP
分析
,
分子植物育种
Vol.9 No.29 (doi: 10.5376/mpb.cn.2011.09.0029)
1212
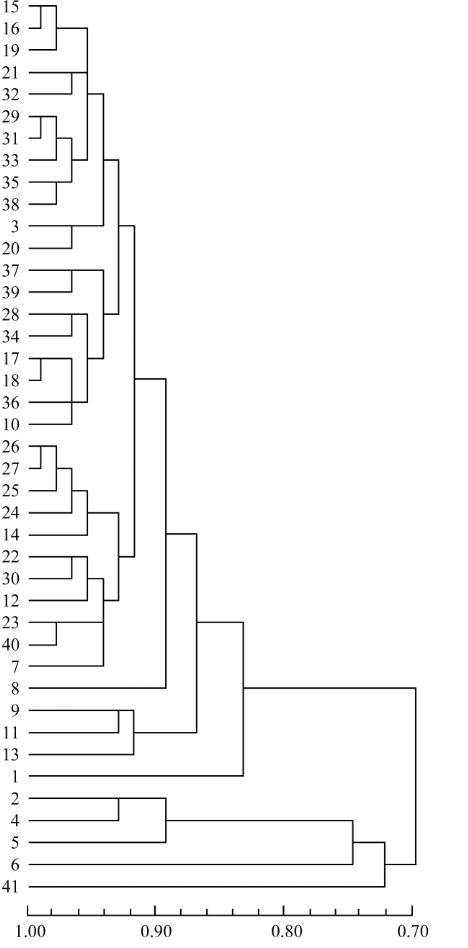
图
2 41
份辣椒材料
SRAP
聚类图
Figure 2 Cluster analysis of 41 accessions in pepper based on
SRAP data
和种间杂交后代
AC3
-
2
-
1 (
种质
7)
、
AC4
-
7
-
3
-
1 (
种
质
12)
;第Ⅱ组为种间杂交材料
AC3
-
6
-
8 (
种质
8)
;
第Ⅲ组由种质
9
、
11
和
13
组成,它们都是源于
C.
annuum
×
C. chinense
种间杂交后代,第Ⅳ组为种间
杂种
B
9431
×H108
。
第Ⅰ组在相似系数
0.921
处进一步分为
2
个亚
组,第
1
亚组有种质
3
、
10
、
15
、
16
、
17
、
18
、
19
、
20
、
21
、
28
、
29
、
31
、
32
、
33
、
34
、
35
、
36
、
37
、
38
和
39
,包含了
C. annuum
中的长椒、圆锥椒和簇生
椒种质,如大果形牛角椒
(
种质
15, 16, 19
和
21)
、早
熟羊角椒
(
种质
3, 29, 31, 32
和
33)
和线椒
(
种质
17, 18
和
20)
等种质都归于此类。第
2
亚组有种质
7
、
12
、
14
、
22
、
23
、
24
、
25
、
26
、
27
、
30
和
40
,包含了
C. annuum
中的灯笼椒、樱桃椒、圆锥椒、长椒和
2
个种间杂
交材料,所有甜椒材料
(
种质
14, 24, 25, 26
和
27)
都归
于此类。
2
讨论
基于
SRAP
技术,本研究对
41
份不同类型辣
椒材料进行了分析,扩增片段的多态性比例高达
46.24%
,材料间遗传相似系数变幅从
0.709~0.982
之间,平均为
0.889
;其中,
C
.
annuum
材料之间遗
传相似系数在
0.923~0.982
之间,平均
0.944
,显著
高于
41
份材料的变幅及总均值,说明辣椒属种间
变异大于种内变异,
C
.
annuum
遗传基础狭窄,这
与基于
RAPD
和
AFLP
技术的研究结果是一致的
(Paran, et al., 1998; Rodriguez, et al. , 1999;
陈学军
等
, 2006)
。
在供试的
29
份
C
.
annuum
材料中,一部分形态
和生物学性状类似的种质首先聚在一起,如种质
14
、
15
和
16
均为大果、中熟牛角椒品种,种质
24
、
25
、
26
和
27
均为甜椒品种。但总的来看,基于
SRAP
标记的分子聚类与
C
.
annuum
形态分类并不对应,
如第
1
小组和第
2
小组均有长椒和圆锥椒材料。其
原因一方面可能是辣椒为常异交自花授粉作物,
C
.
annuum
不同亚种及不同品种之间容易因天然杂交
而产生基因转移现象;另一方面,人工引种打破了
地理隔离的界限,强大的人工选择压力,使品种间
基因转移和交换过程加快,而一些稀有变异类型也
可能在品种改良过程中丢失。
海南野生灌木辣椒植株高大、叶阔卵形,花冠
绿白色,果实单生或双生,果顶直立向上,纺锤状,
晚熟,味香辣,当地素有采食习惯。本研究结果显
示:海南野生灌木辣椒具有美洲同类种质所没有的
特异条带,它们之间的遗传相似系数平均为
0.893
,
远低于
C
.
annuum
材料间的遗传相似系数,说明
海南野生灌木辣椒与美洲灌木辣椒存在较大的遗
传差异。此外,海南野生灌木辣椒与
C
.
annuum
遗传相似系数平均为
0.732
,要小于美洲灌木辣椒