International Journal of Marine Science, 2017, Vol.7, No.33, 316-343
322
1.2.2 Adult aggregations
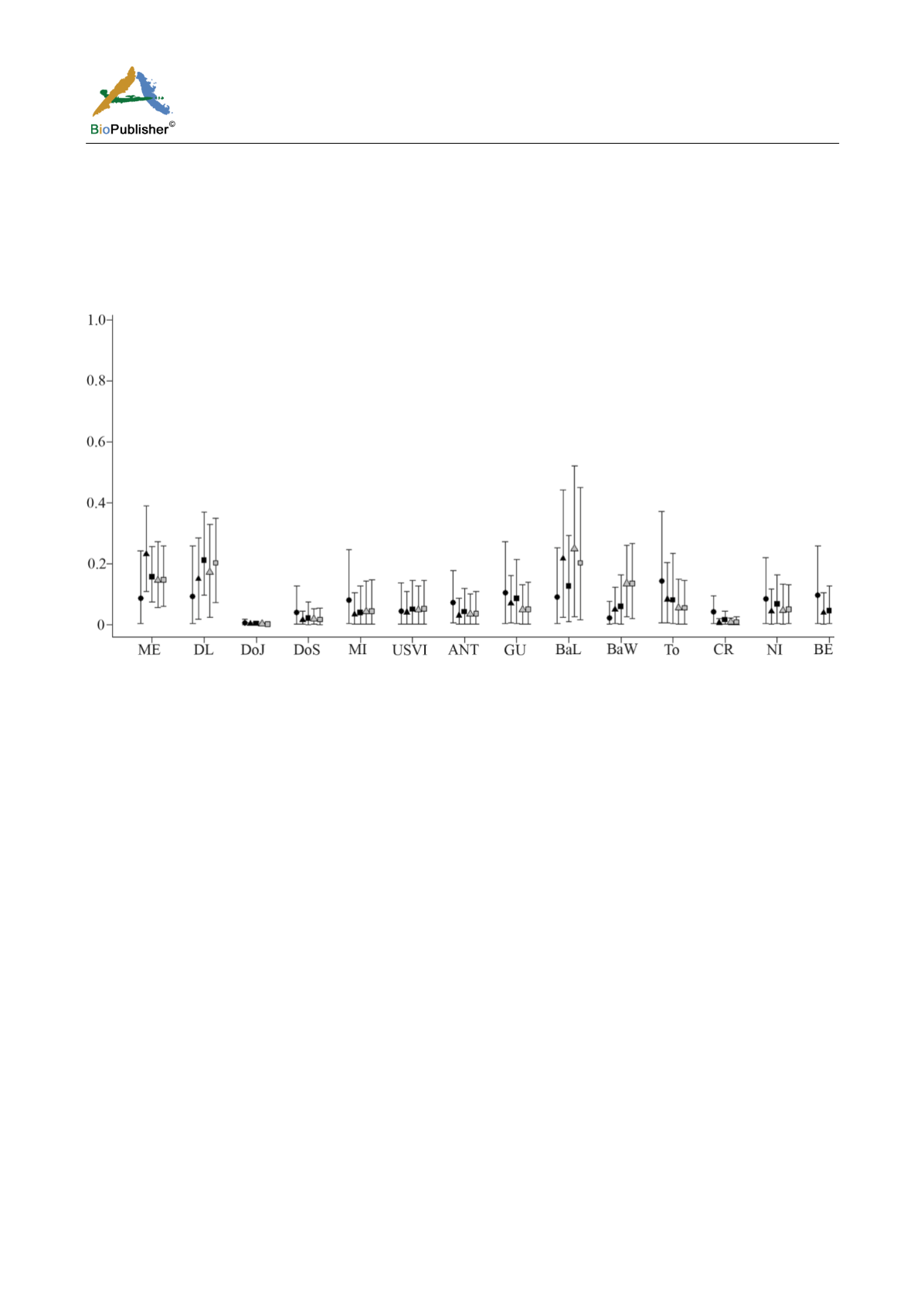
The contributions using the 384 bp length to the adult fishing aggregation of JR were different over the decades
and with respect to the MSAgr (Figure 3). In the 1990s, many rookeries contributed heterogeneously and with low
percentages to this aggregation. In the 2000s, the main contributors were the ME and BaL rookeries followed by
the Cuban rookery; while in MSAgr DL was the major contributor followed by the ME and BaL rookeries. In
contrast, contributions with the 740 bp length haplotypes were similar to MSAgr: BaL as the primary contributor
followed by the DL rookery.
Figure 3 Mixed stock analysis (foraging ground-centric) results showing mean proportional contribution ±2.5–97.5% confidence
intervals for the adult fishing aggregation of JR (black: 384 bp, gray: 740 bp)
Note: Circle and triangles: MSAoa from Díaz-Fernández (1999) and the 2004-2006 fishing period respectively, squares: MSAgr. ME:
Las Coloradas, Yucatan, Mexico; DL: Doce Leguas, Cuba; DoJ: Jaragua National Park, Dominican Republic; DoS: Saona Island,
Dominican Republic; MI: Mona Island, Puerto Rico; USVI: Buck Island, United States Virgin Islands; ANT: Jumby Bay, Antigua;
GU: Trois Ilets, Guadeloupe; BaL: Barbados leeward coast; BaW: Barbados windward coast; To: Tobago; CR: Tortuguero, Costa
Rica; NI: Pearl Cays, Nicaragua; BE: Gales Point, Belize
In Mona Island aggregation of breeding males, the homonymous rookery was the major contributor in each
decade and sequence length (Supplementary Material 1H). In the Doce Leguas fishing aggregation, BaL was the
major contributor during the decade of 1990 followed by the Tobago rookery, while BaL was the dominant
rookery in the MSAgr (Supplementary Material 1H). The Cocodrilo fishing aggregation had heterogeneous
contributions during the 1990s and the MSAgr (Supplementary Material 1I).
The correlations among contributions calculated with both sequence lengths and maritime distances between
rookeries and aggregations were not significant and these had very low coefficients (Table 3).
2 Discussion
When the Cuban Ministry of Fishery reduced the sea turtle harvest by Cuban fishing platforms in the 1990s, JR
and Cocodrilo were the only sites that remained active for reasons of tradition (Carrillo et al., 1999). The
sustainability of this fishery was supported on a supposed recruitment from Doce Leguas rookery to the Cuban
fishing aggregations (Díaz-Fernández et al., 1999), and a fishery minimum size (SCL> 65 cm) based on the fact
that
E. imbricata
individuals of the Cuban platform begin sexual maturation with SCL=60 cm (Moncada and
Nodarse, 1994). Having the original data of Díaz-Fernández (1999) and Lee-González et al. (2015), we could
detect non adult individuals within the harvested hawksbills in both sites. This demonstrates that the mature
cohorts were non accurately managed in the fishery, being these ones well represented in the SCL classes III and
IV and genetically heterogeneous. This places JR in the WC as the only genetically characterized
E. imbricata
fishery aggregation located in a confirmed migratory corridor. Although it is impossible to calculate or estimate


