分子植物育种
(
网络版
), 2016
年
,
第
14
卷
,
第
1024
-
1028
页
Fenzi Zhiwu Yuzhong (Online), 2016, Vol.14, 1024
-
1028
Copyright © 2016 BioPublisher 1025
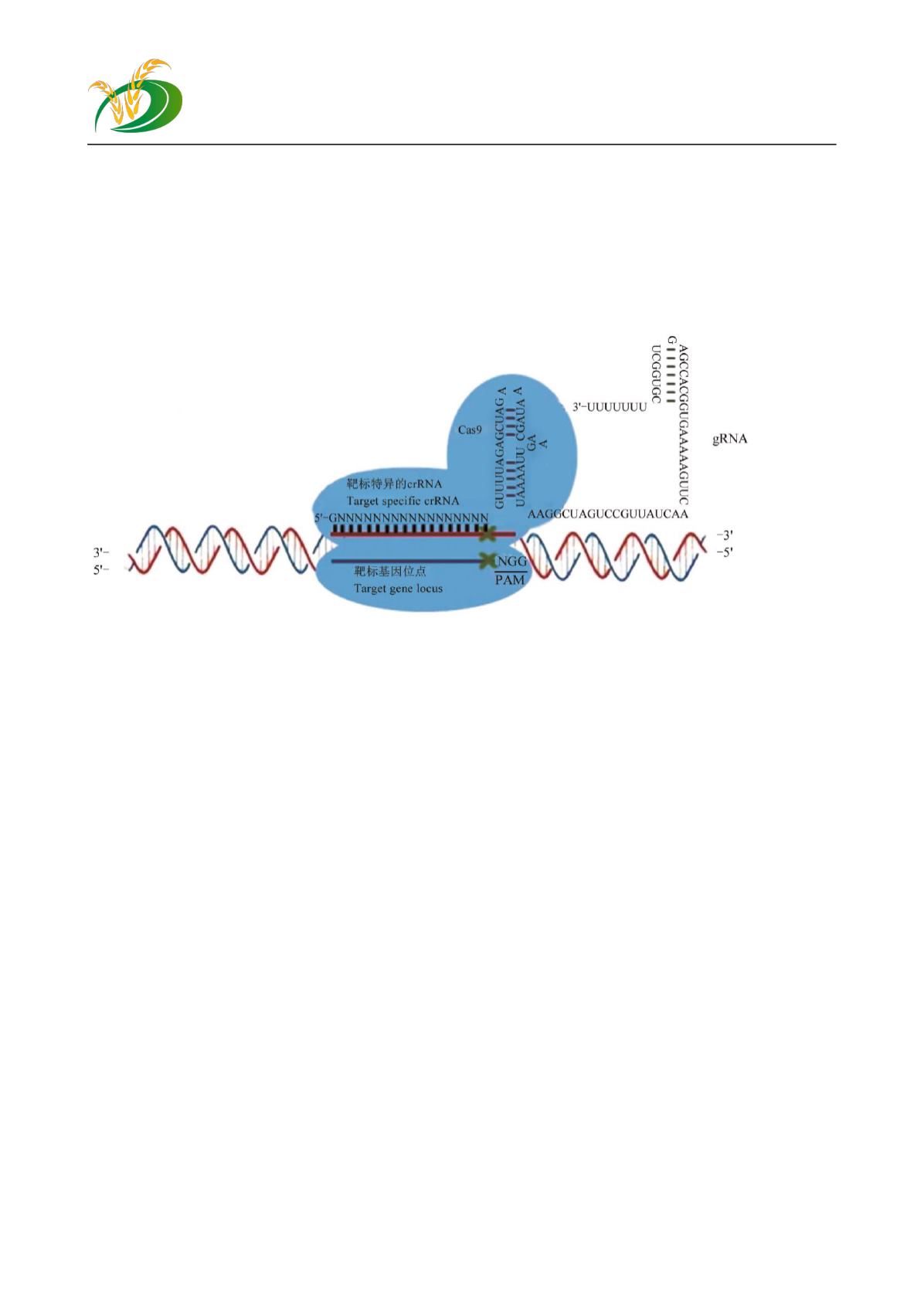
来整合到宿主基因组的
CRISPR
位点的
5′
端;第
二阶段,
CRISPR
基因座的表达,这些插入的短间
隔序列被转录成
crRNA
;第三阶段,
CRISPR-Cas9
系统发挥活性,对外源遗传物质进行干扰。在
Cas
蛋白复合物的参与下,靶向和干扰侵入的噬菌体
DNA
序列
(
李铁民和杜波
, 2011;
方锐等
, 2013)
。
AG
基因是最早克隆的花发育调控基因,与植
物花器官
(
心皮
,
胚珠
,
果实等
)
的发育有密切关
系。由于
AG
基因与植物性别分化,开花结果等
密切相关,因此研究
AG
的调控机制有一定的实
际应用价值。本研究以拟南芥为材料,构建
CRISPR-Cas9
敲除拟南芥
AG
基因载体,建立稳
定的敲除拟南芥基因的转化体系,并获得基因敲
除的突变体。
图
1 CRISPR-Cas9
工作原理示意图
Figure 1 Schematic diagram of CRISPR-Cas9
1
结果与分析
首先将靶位序列片段与克隆载体
pUC19-U3
连
接,转化大肠杆菌,菌落
PCR
筛选阳性克隆
(
图
2)
,
提取质粒为构建好的克隆载体
pUC19-AG
。中间克
隆载体
pUC19-AG
利用
EcoR
I
和
Xba
I
双酶切回收
520 bp
左右片段
(
图
3)
,表达载体
pCAMBIA1302-
Cas9
利用
EcoR
I
和
Nhe
I
分别酶切后过滤小片段并
线性化。利用
T4DNA
连接酶连接
520 bp
片段和线
性化表达载体,构建
pCAMBIA1302-AG
表达载体。
菌落
PCR
筛选引物为
AG-P
:
5’-GCAAATC
GTCTTCTCTAGCCGTGG-3’
和
OsU3-D:CGGCTA
GAGAAGACGATTTG
。
表达载体构建过程中
pCR
筛选验证的电泳图说
明表达载体已构建成功,并可以进行转入植物体内
进行表达过程。
2
讨论
虽然
CRISPR
系统的发现可以追溯到
1987
年,
但是
CRISPR-Cas9
这种技术的应用研究还处于初始
阶段,是近几年才逐渐发展起来的。该技术已经成
功应用在多种微生物、动物细胞、动物和植物。
2013
年
Nature Biotechnology
同时报道了在重要作物水
稻、小麦以及模式植物拟南芥和本生烟中取得了多
个基因定点敲除、插入等基因组定点编辑的操作的
成功案例,并首次证实
CRISPR-Cas9
系统能够在植
物基因组中实现定点编辑
(Shan et al., 2013; Feng et
al., 2013; Li et al., 2013; Nekrasov et al., 2013)
。
利用
CRISPR-Cas9
系统极性基因定点编辑可
以用更加灵活性的方式实现只利用载体定点编辑同
一个基因的不同位点和利用一个载体同时编辑多个
靶位点。由于此系统的靶位点很短,只有
20 bp
左
右,所以可以设计几个串联在一起的
sgRNA
,来实
现对同一基因多个位点的同时编辑以及多个不同基
因靶位点的编辑。同时定点编辑多个基因,可以有
助于研究基因间的相互作用机制和研究同一基因家
族的不同基因的功能。
CRISPR-Cas9
在各项研究领域必将有越来越
有广阔的应用前景,为基因组定向编辑领域的研究
带来突破性的技术革命
,
特别是在基因功能解析和
加速重要农作物水稻、小麦性状改良与分子定向育
种等方面
(
李君等
, 2013)
。


