分子植物育种
(
网络版
), 2016
年
,
第
14
卷
,
第
1087-1094
页
Fenzi Zhiwu Yuzhong (Online), 2016, Vol.14, 1087-1094
Copyright © 2016 BioPublisher 1090
1.3.2 Cdc48E
,
Per12
和
Hsp17.6
等低温萌发相关响
应蛋白的表达量与低温萌发关系的分析
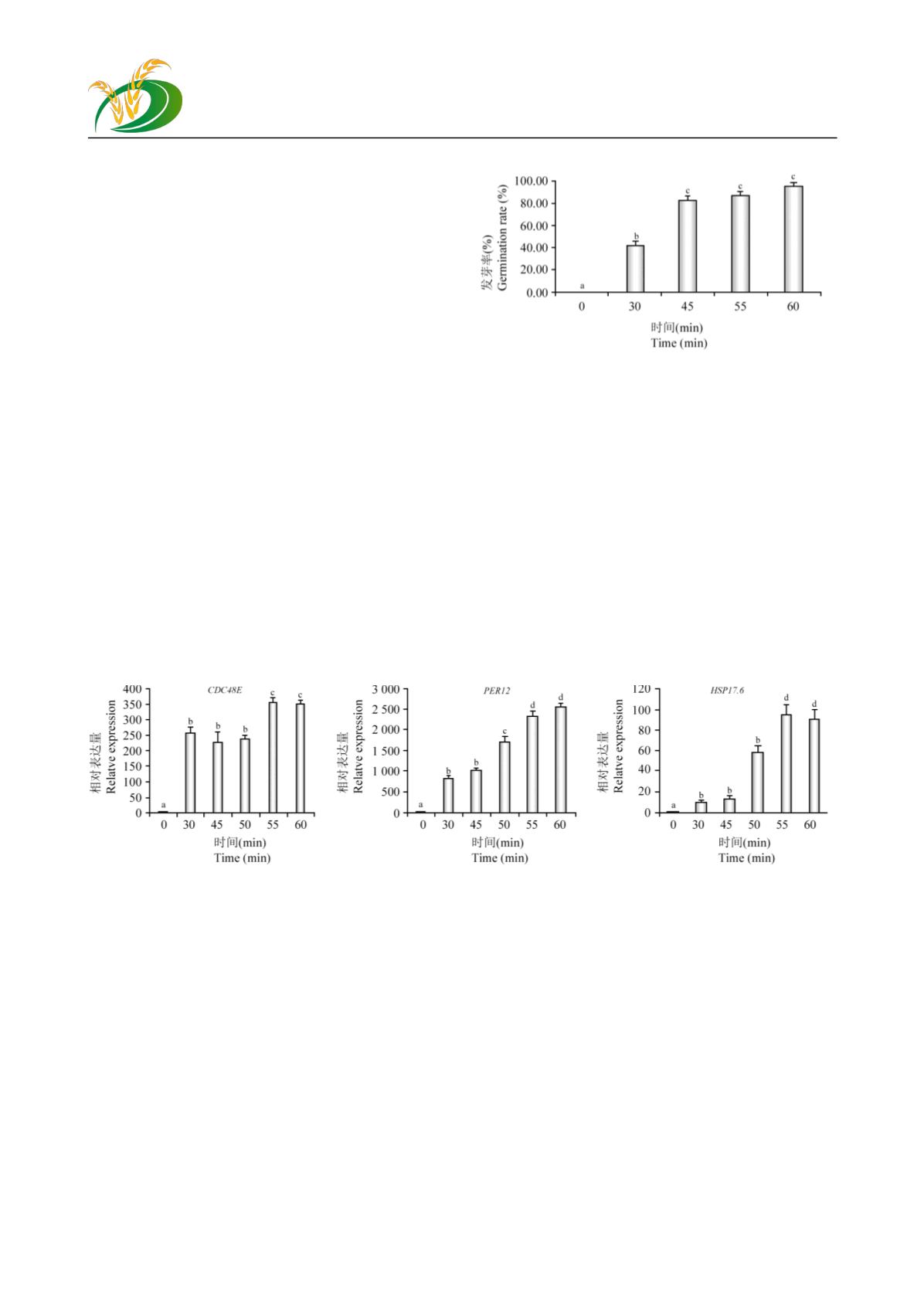
独行菜种子在
4
℃下层积
10 d
后,处于萌发停
滞期。用
25
℃分别处理
30 min
,
45 min
,
55 min
,
1 h
,统计不同时间处理对独行菜种子耐受低温萌发
率的影响。经
25
℃,
30 min
,
45 min
,
55 min
,
1 h
诱导处理处于萌发停滞期的独行菜种子后,在
4
℃
条件下萌发率分别达到
0%
、
40%
,
83.33%
,
88.33%
,
96.11%
;也就是经
30 min
的
25
℃处理低
温停滞期的种子后,
40%
的种子解除萌发停滞,耐
受低温继续进行萌发;经
45 min
的
25
℃处理,
83.33%
的种子解除萌发停滞;经
1 h
的
25
℃处理,
种子耐受低温萌发率达到正常温度萌发率,也就是
几乎所有的种子能够解除萌发停滞
(
图
3)
。
为进一步探讨筛选获得的相关响应蛋白表达
与独行菜低温萌发间的关系,对
CDC48E
,
PER12
和
HSP17.6
三个基因在经不同时间
25
℃诱导处理
处于萌发低温停滞期的种子中的表达情况进行分
析。
25
℃处理
30 min
后,与处于低温停滞期的独行
菜种子相比较,
CDC48E
,
PER12
和
HSP17.6
基因
的表达量均极显著提高
(p<0.01)
,且在
1 h
内随处理
图
3 25
℃处理不同时间对独行菜种子低温萌发的影响
Figure 3 Germination rate of the
Lepidium
seeds under low
temperature after dealing with 25
℃
for different time
时间的增加均呈上升趋势
(
图
4)
。三个基因的表达特
性也有一定的差异,其中
CDC48E
、
PER12
基因经
25
℃处理不同时间均有极显著上调;而
HSP17.6
基
因表达量在
25
℃处理
50~60 min
时极显著上调
(p<0.01)
,而在
25
℃处理
45 min
以下显著上调表达
(p<0.05)
。这与经
25
℃处理超过
45 min
后,
80%
以
上的种子都能解除萌发停滞期,耐受低温继续进行
萌发的变化趋势一致。这也说明筛选获得的
25
℃诱
导上调表达的蛋白对独行菜种子低温萌发停滞解
除关系密切。
图
4 25
℃处理不同时间后独行菜种子低温萌发相关基因的
qRT-PCR
Figure 4 The genes expression analysis at the different germination stage of
Lepidium apetalum
Willd.
2
讨论
种子萌发是一个非常复杂的过程,涉及到许多
与生长发育相关的蛋白的调控表达。本研究以生活
在新疆北部的独行菜种子这一特殊生态型为研究
材料,通过比较其低温萌发停滞解除前后蛋白表达
的差异,筛选出停滞解除表达量显著上调的蛋白并
进行质谱鉴定分析,并对其中部分蛋白做了相应的
基因表达分析。
热激蛋白是一类在系统发育过程中高度保守
的分子伴侣,按相对分子量
(KD)
大小可以分为
Hsp100s
,
Hsp90s
,
Hsp70s
,
Hsp60s
和
sHsps
。前四
种
Hsp
被视为
HWM Hsps (high molecular weight
Hsps)
,它们主要参与一些蛋白的折叠和装配。
Hsp70B
定位在细胞质膜,叶绿体和细胞质中,与
其它分子伴侣协同作用,参与胞质溶胶和细胞器中
新生蛋白的富集和折叠是热激诱导合成的,能减轻
植物在冷环境中的膜伤害,增强组织的耐冷性,与
植物获得耐受性关系密切
(Lin et al., 2001)
,黄上志
等发现萌发的水稻种子经
42
℃热激处理后其幼苗
的耐冷性明显增强
(
黄上志等
, 2004)
。细胞质中分子
陪伴蛋白
Cpn60-α1
亚基可以结合
Rubisco
大亚基
(
陈根云
, 2000)
,帮助蛋白正确折叠,免受其他蛋白


