Basic HTML Version









分子植物育种
(
网络版
), 2012
年
,
第
10
卷
,
第
1376
-
1382
页
Fenzi Zhiwu Yuzhong (Online), 2012, Vol.10, 1376
-
1382
http://mpb.5th.sophiapublisher.com
1379
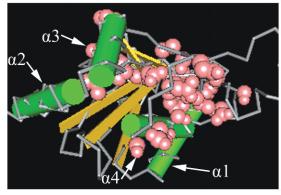
图
3
油茶
UBE2S
泛素结合酶催化位点
(UBCc)
分析
注
: A:
比对的序列
; B:
半胱氨酸活性位点
; C:
与泛素形成硫酯键中间产物的残基
; D:
与
E3
相互作用的残基
; E:
特异性注释
;
F:
超级家族
Figure 3 Analysis of ubiquitin-conjugating enzyme E2, catalytic(UBCc) domain in
Camellia oleifera
UBE2S
Note: A: Query sequence; B: Active site cysteine; C: Ub thioester intermediate interaction residues; D: E3 interaction residues; E:
Specific hits; F: Superfamilies
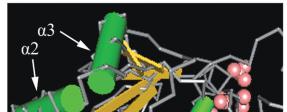
图
4
位于
94
位的半胱氨酸残基在保守区域中的位置
Figure 4 Active site cysteine on conserved domain UBCc
图
5
与泛素通过硫酯键结合的残基在保守区域中的位置
Figure 5 Ub thioester intermediate interaction residues on
conserved domain UBCc
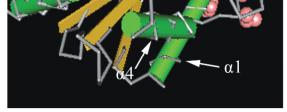
图
6
与
E3
相互作用的残基位点在保守区域的位置
Figure 6 E3 interaction residues on conserved domain UBCc
2
讨论
泛素
/26S
蛋白酶体途径能够选择性的降解蛋
白质,这在许多代谢过程中起关键作用,是真核生
物重要的翻译后修饰系统,如参与调控细胞周期、应
急反应、信号传导及
DNA
修复等
(
陈建明和余应年
,
2000)
。蛋白质发生单泛素化不会水解,但可调节其
生物学活性,如调节蛋白质稳定性、蛋白三级结构、
改变染色体结构、基因修复、复制及表达调节等。
靶蛋白的多泛素化需要进行多轮单泛素化反
应,每轮单泛素化过程均是一系列的酶促级联反
应,
E2
在单泛素化过程中催化第二步反应。目前研
究表明,所有真核生物都有多种
E2
,例如,已发现
人类的
E2
至少有
30
种,拟南芥有
37
种,不同的
E2
在对靶蛋白的特异性选择上具有重要作用。推测
正是由于每个物种中均有一个庞大的
E2
基因家族,
大多数物种中已克隆的
E2
基因数量较少,因此油
茶
UBE2S
基因在
genebank
中只比对到
2
个同源性
较高的序列。
E2
根据其结构和功能大致可以分为
4
类
(
陈建明和余应年
, 2000; Plafker et al., 2004; Ma-
tuschewski et al., 1966; Jentsch, 1992)
:Ⅰ类
E2
,只
有一个保守催化结构域
(UBC)
,长约
150 AA
,其功
能主要是参与短周期蛋白和异常蛋白的泛素依赖
性降解;Ⅱ类
E2
,由
UBC
和一个
C
-末端延伸结构
组成,其
C
-末端结构可参与识别目标蛋白、与
E3
酶相互作用等;Ⅲ类
E2
,由
UBC
和一个
N
-末端延
伸结构组成,目前尚不清楚
N
-末端结构的功能;
IV
类
E2
,同时具备
UBC
、
C
-末端结构和
N
-末端结构。
油茶
UBE2S
含有
E2
的保守催化结构域、活性位点
半胱氨酸残基,第
146
位
~270
位
AA
为
C
-末端延