Basic HTML Version





分子植物育种
(
网络版
), 2012
年
,
第
10
卷
,
第
1067
-
1079
页
Fenzi Zhiwu Yuzhong (Online), 2012, Vol.10, 1067
-
1079
http://mpb.5th.sophiapublisher.com
1068
来对拟南芥和水稻的大量卓有成效的研究,为完整
地揭示植物生殖细胞分化和配子体形成的分子机理
增添了丰富内容。本文正是基于拟南芥和水稻等植
物的雌配子体发育的分子遗传研究的最新进展,重
点阐述雌配子体发育的分子机理和遗传调控过程。
植物雌配子体的发生涉及一系列细胞学的演
化过程,其中包括孢原细胞
(archesporial cell, AC)
的
形成、大孢子母细胞
(megaspore mother cell, MMC)
分化、
MMC
减数分裂、功能大孢子的确立、胚囊
(Embryo sac)
的形成、有丝分裂和胚囊极性化形成成
熟胚囊等
(
图
1)
。以高等植物蓼科
(Polygonaceae)
为
例,雌配子体的细胞结构一般包含
2
个助细胞
(synergid cell)
、
1
个卵细胞
(egg cell)
、
1
个中央细胞
(centre cell)
、
3
个反足细胞
(antipodal cell)
,是一个
具有八个细胞核的七细胞的胚囊结构,拟南芥和水
稻的雌配子体属于蓼型胚囊
(
polygonum type
)
。
MMC
减数分裂是植物雌配子体的发育的一个极
其重要的标志性事件,因此,生殖发育研究中往
往以
MMC
减数分裂为界,其前的细胞学过程为
孢子体形成
(sporogenesis)
,其后的为配子体形成
(gametogenesis)
。本文以其为界,分别叙述两个发
育过程的相关研究。
1
参与植物孢子体形成
(sporogenesis)
相关基因
1.1
调控孢原细胞发生与大孢子母细胞发育的基因
拟南芥和水稻等植物的珠心中表皮下一个
L2
细胞开始分化,其接近等径、细胞核大、胞质丰富,
明显有别于其他珠心细胞,这个细胞称为孢原细胞
(archesporial cell, AC)
。孢原细胞伸长变大,形成大
孢子母细胞
(megasporocyte cell, MC)
,随后进入减
数分裂期。
在雌配子体发生过程中,体细胞是如何启动并
分化形成生殖细胞或孢原细胞是学者感兴趣的
问题之一。通过对拟南芥突变体研究,发现
SPOROCYTELESS
(
SPL
)
/NOZZLE
(
NZZ
)
基因可能参
与了这一过程
(Schiefthaler et al., 1999; Yang et al.,
1999)
。在
spl/nzz
突变体中,孢原细胞的起始正常,
然而随后的大孢子母细胞和小孢子母细胞分化受
阻,其花粉囊壁发育也未发生,尽管珠心发育延迟
了但是珠被发育正常
(Yang et al., 1999)
。当
SPL
过
表达时,可以使拟南芥的花瓣体细胞变成生殖细
胞,经小孢子发生途径直接形成花粉
(Ito et al.,
2004)
。这表明,
SPL/NZZ
与周围细胞的相互作用决
定了花药和胚珠的产生。突变体
sterile apetela
(
sap
)
的大孢子母细胞不能完成减数分裂,小花的数目和
器官的数目也受到影响
(Byzova et al., 1999)
。然而,
在突变体
sap
中大孢子母细胞已形成,因此认为
SAP
可能作用于
SPL/NZZ
下游。
由于玉米的
MULTIPLE ARCHESPORIAL CELLS1
(
MAC1
)
基因可以改变孢原细胞的定向分化,因此认
为
MAC1
基因在孢原细胞发育中可能起作用时期比
SPL/NZZ
基因更早。在
mac1
突变体中,其胚珠中
形成了多个孢原细胞
(Sheridan et al., 1996)
。但是,
其花药的孢原细胞定向分化并没有受到影响,而是
在减数分裂时被中止,这暗示了在花药的发育过程
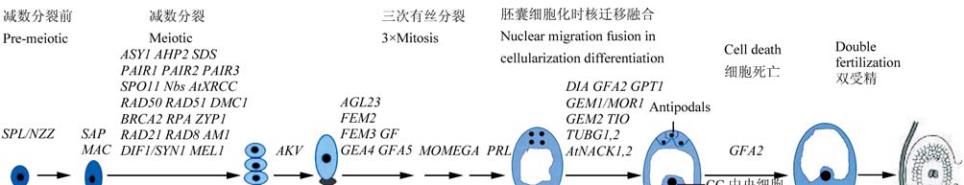
图
1
植物雌配子体发育过程中孢子发生和配子发生的连续过程及参与的相关基因示意图
Figure 1 Schematic diagram depicting the sequential processes of sporogenesis and gametogenesis in female gametophyte
developement with related genes in plant