Basic HTML Version




分子植物育种
(
网络版
), 2012
年
,
第
10
卷
,
第
1061
-
1066
页
Fenzi Zhiwu Yuzhong (Online), 2012, Vol.10, 1061
-
1066
http://mpb.5th.sophiapublisher.com
1062
境破坏和高价位的兰花使得来源于农村广袤无垠
的林地的国兰资源变得极度濒危或十分稀有
(
吴应
祥
, 1993;
江泽慧
, 2000)
,因此,探索奇花形成的分
子机理等遗传学基础研究,对保护国兰野生资源,
培育优良国兰品种具有重要意义。
在兰花中蝶化程度的高低是区别蝶瓣花品质
优劣的主要依据,
‘
苍山奇蝶
’
为莲瓣兰
(
Cymbidium
tortisepalum
)
蝶瓣奇花,
1990
年始发现于云南省大理
州境内。此品种捧瓣蝶化面积达
90%
以上,具有重
要的观赏价值和经济价值
(
刘振龙
, 2007)
。
研究人员在对拟南芥和金鱼草的各类花器官
同形突变体的研究中,发现了在花的发育中调控各
类花器官形成的
A
、
B
、
C
、
D
、
E
基因。这几类基因
中的一类或几类发生突变就会导致萼片变成花瓣,
花瓣变成雄蕊等
(
朱永平等
, 2011)
。这些基因称为同
形异位基因,它们编码的蛋白质均含有一个保守性
很强的
DNA
结合序列
——MADS
盒,因此属于
MADS
盒基因家族。该家族每一类基因通过基因复
制而产生亚家族
,
而后又独自地发生了基因复制事
件,产生多个平行进化同源基因
(Coen et al., 1991;
Pelaz et al., 2000)
。
现有的研究揭示了
MADS-box
等基因在兰花成
花转换及花器官发育过程中的重要作用
(Lu et al.,
1993; Mariana et al., 2008)
。近年来,关于兰科植物
开花调控的研究取得了一定的进展,已分离鉴定出
一些花发育调控基因,包括花器官特异基因及一些
花分生组织特异基因,但主要集中在热带兰上,而
对中国兰的研究少有报道。对于蝶化奇花基因的研
究几乎没有报道。
本试验利用消减抑制杂交技术对
‘
苍山奇蝶
’
蝶
化捧瓣与其正格花捧瓣之间的差异表达基因进行
研究,以期探索中国兰花花型瓣化变异机理,为利
用植物基因工程改良中国兰花的观赏性状提供理
论依据。
1
结果与分析
1.1
总
RNA
及
mRNA
的质量检测
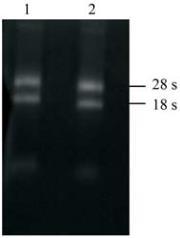
提取
‘
苍山奇蝶
’
蝶化捧瓣和正格花捧瓣的总
RNA
后,在紫外呈像仪上观察
RNA
条带。结果显示
(
图
1)
总
RNA
具有
28S
、
18S
两条明显的带谱,其完整
性较好,且
28S
和
18S
的亮度比例约为
2:1
,表明提取
的总
RNA
可用构建文库。
1.2
酶切效果分析
电泳结果图
2A
表示蝶化捧瓣合成
cDNA
的片段
图
1
莲瓣兰总
RNA
注
: 1:
蝶化捧瓣
; 2:
正格花捧瓣
Figure 1 Identification of total RNA in
Cymbidium tortisepalum
Note: 1: Lip-like petals; 2: Normal petals
大小分布与
8 000~100 bp
之间,经
Rsa
Ⅰ酶切后,条
带明显下移,集中于
100~1 000 bp
之间,图
2B
表示
正格花捧瓣合成
cDNA
的片段大小分布与
5 000~
500 bp
之间,经
Rsa
Ⅰ酶切后,条带明显下移,集中
于
100~1 000 bp
之间,大片段减少、小片段增多。
说明酶切效果好,适用于后续试验。
图
2
蝶化捧瓣
(A)
及正格花捧瓣
(B)
的酶切和未酶切
cDNA
注
: M: 8 kb DNA marker; 1:
未酶切
; 2:
酶切
Figure 2 The products of undigested and digested cDNA in
lip-like petals (A) and normal petals (B)
Note: M: 8 kb DNA marker; 1: Undigested; 2: Digested
1
.3
抑制消减
cDNA
文库中
cDNA
片段大小的检测
构建的正向文库大小为
3 000
个克隆,反向文库
大小为
500
个克隆。从正向文库中随机挑选
15
个菌
落进行菌落
PCR
,外源片段的长度分布在
100~750
bp
左右,平均片段长度约
500 bp
左右
(
图
3)
。以上结
果显示,成功构建消减抑制文库。
1.4 ESTs
功能分类
将唯一序列
NCBI
在线
Blastx
进行同源比对及
功能分析。
114
个序列按功能分类为
MADS
、假设蛋
白、
GenBank
中未匹配蛋白、储存蛋白以及其它等
5
类
(
图
4)
。其中
MADS
基因所占比例为
5%
、假设蛋白