Basic HTML Version




分子植物育种
(
网络版
), 2012
年
,
第
10
卷
,
第
1479-1486
页
Fenzi Zhiwu Yuzhong (Online), 2012, Vol.10, 1479-1486
http://mpb.5th.sophiapublisher.com
1482
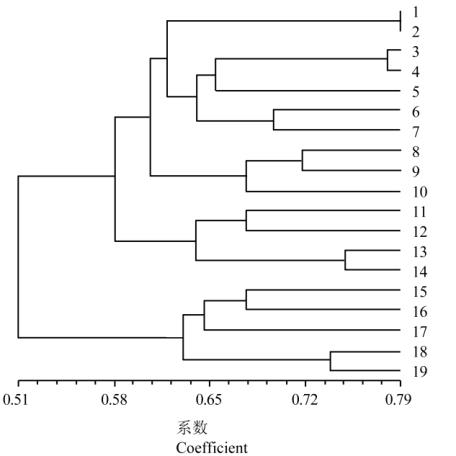
群可明确区分国外引进的品种与国内品种,表明品
种间的亲缘关系较远,受地理分布影响较大,国内
啤酒大麦品种与国外抗赤霉病大麦品种的种质渗
透较少。以相似系数
0.61
为阈值,来源于国内品种
的第一类群可以分为
3
个亚类群。第一亚类群和第
三亚类群分别为抗扩展性较好大麦品种或育种材
料;第二类群包括连
9723
、单二和甘啤
2
号,为赤
霉病抗性较差的
3
个中国大麦品种。第二大类群中,
六棱大麦和二棱大麦分别位于两个亚类群中。
图
1
大麦品种基于
SSR
标记树状聚类图
注
: 1:
扬农啤
5
号
; 2:
苏花
2
号
; 3:
新宇
03~3; 4:
苏
B0608; 5:
申河
085; 6:
扬辐
6008; 7:
如东
5218; 8:
连
9723; 9:
单二
; 10:
甘啤
2
号
; 11:
苏啤
4
号
; 12:
苏啤
3
号
; 13: 5E003; 14:
盐
99~175; 15: Cheveron; 16: Morrison; 17: Vivar; 18: CIho4196;
19: Frederickson
Figure 1 The dendrogram constructed based on polymorphic
SSR data in 19 barley varieties
Note: 1: Yangnongpi5; 2: Suhua2; 3: Xinyu03~3; 4: SuB0608;
5: Shenhe085; 6: Yangfu6008; 7: Rudong5218; 8: Lian9723 9:
Daner; 10: Ganpi2; 11: Supi4; 12: Supi3; 13: 5E003; 14:
Yan99~175; 15: Cheveron; 16: Morrison; 17: Vivar; 18:
CIho4196; 19: Frederickson
1.3
大麦品种赤霉病抗性基因的等位位点变异
据前人报道,在
2H
染色体上存在
QTL1
、
QTL2
、
QTL3
等
3
个赤霉病抗性
QTL
,在
3H
染色体上存在抗
赤霉病
QTL4
,在
6H
染色体上存在抗赤霉病
QTL5
。
以供试品种的抗性
QTL
紧密连锁的分子标记的多
态性位点差异来分析抗性基因可能的异同,由表
2
可见,在供试的抗赤霉病啤酒大麦品种中,新宇
03-3
与连
9723
可能携带
QTL1
、
QTL2
和
QTL3
等
3
个
QTL
位点,苏花
2
号可能携带
QTL2
和
QTL3
,苏
B0608
可
能携带
QTL1
和
QTL2
,扬农啤
5
号可能携带
QTL2
和
QTL3
,苏啤
3
号可能携带
QTL1
和
QTL3
,
5E003
、
苏啤
4
号和如东
5218
可能携带
QTL3
,申河
085
和扬
辐
6008
可能携带
QTL2
。盐
99175
虽然具有较好的抗
性,但并未检测到已报道的
QTL
,而中感品种单二
和感病品种甘啤
2
号却检测到可能携带有
QTL1
和
QTL2
。上述结果提示这些已报道的抗性基因间并
非是简单的加性效应,品种的抗病表现与已报道的
抗性位点难以构成直接的相关关系,可能在抗病品
种中仍然存在其他抗病基因。感病品种中携带与抗
病
QTL
连锁标记一致的等位位点,说明抗病基因与
感病基因可能在同一品种中是同时存在的。
2
讨论
前人研究已经明确,小麦赤霉病抗性存在抗初
侵染和抗扩展
2
种抗病类型
(Schroeder and Chris-
tensen, 1963)
。但有关大麦赤霉病抗性,对这
2
类抗
性的则看法不一,早期研究认为大麦仅存在抗侵
染,不存在抗扩展,病症不会由发病小穗向邻近小
穗扩展
(Steffenson, 2003; Bai and Shaner, 2004)
。但
Bushnell
等
(2003)
的研究证实大麦存在抗扩展性,试
验以单花滴注方式评价了大麦品种的抗扩展性
,
发
现供试品种间存在明显的抗扩展性差异
,
并在大麦
2H
染色体上检测出
1
个抗扩展性
QTL
。国内的研究
也证实,大麦赤霉病菌侵染寄主后存在明显的扩展
过程,不同大麦品种的赤霉病对赤霉病菌的扩展抗
性存在显著差异
(
戈和静等
, 2006;
吴佳祺等
, 2011)
。
采用单花滴注方式接种并对接种后赤霉病小穗数
或病小穗率是进行鉴定可明确区分赤霉病抗扩展
性,利用孢子喷雾接种方式并统计接种后的病小穗
率或病小穗数可作为抗初侵染的鉴定方法
(Bai and
Shaner, 2004)
。本试验采用了上述两种方法进行接
种,试图区别抗扩展和抗侵染
2
种抗性类型,但从
单花滴注接种后
21 d
的病小穗率与孢子喷雾接种后
14 d
的病小穗数的相关性看,品种间未表现出明显
的抗性差异,无法区分抗侵染和抗扩展
,
主要原因可
能由于在接种后
14 d
进行调查,病小穗数的差异受
到了抗扩展能力的影响。从育种实际出发,单一研
究抗侵染的意义并不重要,在病原菌侵染后其产生
的菌丝总是在不断扩展的,最终的病小穗数才真正
反应出品种的抗病性。试验还调查了孢子喷雾接种