Basic HTML Version




分子植物育种
(
网络版
), 2012
年
,
第
10
卷
,
第
1278
-
1286
页
Fenzi Zhiwu Yuzhong (Online), 2012, Vol.10, 1278
-
1286
http://mpb.5th.sophiapublisher.com
1281
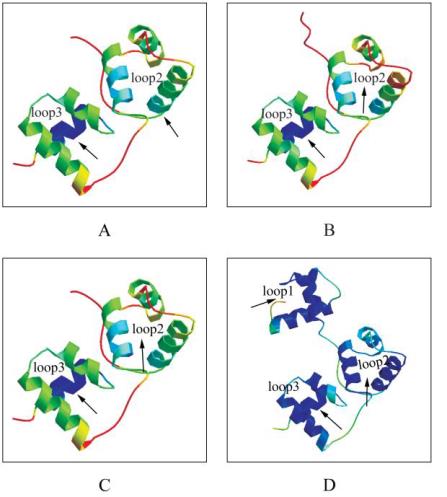
图
3
Cl
MYB4
蛋白的三维空间结构预测及与
At
MYB46
和
Pt
MYB46
蛋白构象比较
注
: A, B, C:
分别代表
Cl
MYB4,
At
MYB46
和
Pt
MYB4
蛋白
R2R3
结构域预测的空间结构
; D:
是已知空间结构蛋白
(1H8CC)
的
R1R2R3
结构域
,
为同源建模的模板空间结构
Figure 3 Predicted three-D structure of
Cl
MYB4 protein and
compared with
At
MYB46 and
Pt
MYB4 proteins
Note: A, B, C: Predicted three-D structures of R2R3-domain
in
Cl
MYB4,
At
MYB46 and
Pt
MYB4 proteins; D: Three-D
structure of 1H8CC as the template
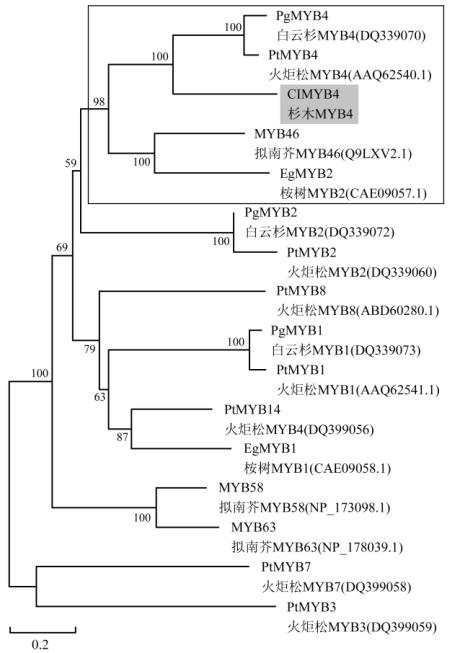
利用
MEGA 5.0
软件的
Neighbor-Joining
法,构
建了基于蛋白氨基酸序列的系统进化树,如图
4
。
所研究的蛋白序列一部分为裸子植物火炬松和白
云杉的
R2R3
-
MYB
基因,一部分为桉树和拟南芥
中与次生壁和木质素合成转录调控相关的
MYB
基
因。结果显示,
Cl
MYB4
与白云杉
Pg
MYB4
,火炬
松
Pt
MYB4
,桉树
Eg
MYB2
和拟南芥
At
MYB46
聚
为一个亚组,拟南芥
At
MYB46
已被证实是维管发
育决定基因
SND1 (Secondary cell wall related NAC
Domain1)
的直接下游靶基因
(Zhong et al., 2007)
,负
责次生壁合成过程的转录调控。其他裸子植物
MYB
基因中,
Pg
MYB2
与
Pt
MYB2
、
Pt
MYB3
与
Pt
MYB7
、
Pt
MYB1
、
Pt
MYB8
和
Pg
MYB1
各自聚为一个小类,
说明
Cl
MYB4
与白云杉
Pg
MYB4
,火炬松
Pt
MYB4
的亲缘关系最近。此外,拟南芥
At
MYB58
和
At
MYB63
单独聚为一类,而桉树
Eg
MYB2
和火炬
松
Pt
MYB14
聚为一类。
图
4
基于氨基酸序列构建的
Cl
MYB4
系统发育树
注
:
方框内为参与次生壁合成调控的
R2R3-MYB
基因蛋白
亚族
Figure 4 Phylogenetic tree based on amino acid sequences of
Cl
MYB4
Note: The box indicated the R2R3-MYB protein subgroup
related to secondary cell wall biosynthesis
1.3
杉木
MYB
基因的原核表达条件优化
将构建正确的原核表达载体质粒
pET
-
30b-
Cl
MYB4
转化大肠杆菌
Rosetta
菌株。挑选阳性克
隆子在
37
℃条件下采用
IPTG
浓度
0.01 mmol/L
,
0.1 mmol/L
,
1 mmol/L
诱导后,
SDS-PAGE
电泳显
示目的蛋白
Cl
MYB4
均可以表达,但是
Cl
MYB4
蛋白
(70 kD,
如图
5)
的大小与预测蛋白分子量大小
(54 kD)
不太一致。不同的
IPTG
对重组蛋白表达的
诱导影响不明显,
0.1 mmol/L
,
1 mmol/L IPTG
诱导
的效果较好,因此,我们选择
0.1 mmol/L IPTG
作
为大量诱导重组蛋白表达的使用浓度。
1.4
杉木
MYB
基因原核表达蛋白的纯化
分析发现
Cl
MYB4
重组蛋白不是以可溶性蛋
白形式存在,而是主要存在于大肠杆菌的包涵体