Basic HTML Version




分子植物育种
(
网络版
), 2012
年
,
第
10
卷
,
第
1259
-
1264
页
Fenzi Zhiwu Yuzhong (Online), 2012, Vol.10, 1259
-
1264
http://mpb.5th.sophiapublisher.com
1262
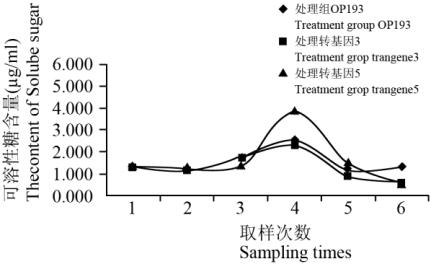
抽穗期干旱胁迫下,对照组和处理组含和不含
CBF3
水稻的可溶性糖含量变化如图
6
、图
7
所示。
从图
6
、图
7
可以看出,对照组和处理组对应材
料间可溶性糖含量大小没有明显差异,其变化趋势
也基本相同,都是先上升再下降,再小幅提高,并
在第
4
次取样时有一个峰值。但不同材料间则存在
差异,对照组:转基因
5 NT
的峰值>含
CBF3
的
OP193
>转基因
3
,干旱处理组:转基因
5 NT
的峰值
>含
CBF3
的
OP193
和转基因
3
,而
OP193
和转基因
3
则几乎相同。由此表明:抽穗期干旱胁迫时水稻可
溶性糖含量的变化与
CBF3
基因的存在无关联。
1.5
抽穗期干旱胁迫下的含和不含
CBF3
水稻的可溶
性蛋白含量变化
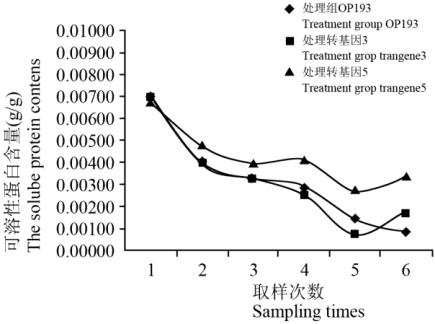
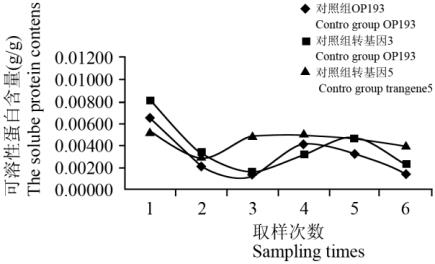
抽穗期干旱胁迫下,对照组和处理组含和不含
CBF3
水稻的可溶性蛋白含量变化如图
8
、图
9
所示。
从图
8
、图
9
可以看出,对照组:
3
个材料可溶
图
7
处理组试验材料可溶性糖含量变化
Figure 7 Variation of content of soluble sugar of the treatment
图
8
处理组可溶性蛋白含量变化
Figure8 variation of content of soluble protein of the treatment
图
9
对照组可溶性蛋白含量变化
Figure 9 variation of content of soluble protein of the control
性蛋白含量变化存在先下降,再上升再下降的波浪
形下降趋势,而干旱处理组对应材料间可溶性蛋白
含量的波浪形下降趋势更明显。不同材料间也存在差
异,对照组:转基因
5 NT
的变化幅度<含
CBF3
的
OP193
<转基因
3
,干旱处理组:转基因
5 NT
的变化幅度<含
CBF3
的
OP193
和转基因
3
,而
OP193
和转基因
3
则几
乎相同。结果表明:抽穗期干旱胁迫时,
CBF3
基因
的存在对水稻可溶性蛋白含量的变化影响不大。
2
讨论
渗透调节是植物防御干旱的一种重要方式,渗
透调节的生理效应是增加细胞溶质浓度,降低渗透
势,保持膨压,缓解脱水胁迫伤害,有利于保持水
分和细胞各生理过程的正常进行
(
白向历
, 2007)
。脯
氨酸、可溶性糖、可溶性蛋白作为植物主要的渗透
调节物质之一,近年来被广泛研究
(
黄文江等
,
2002)
。叶绿素是植物最重要的收集转换光能的光受
体色素
,
叶片失绿被认为是叶片衰老的最主要和最
初的症状
(
孙耀中和东方阳
, 2008)
。在干旱胁迫下,
叶片叶绿素含量越高、
Chla/Chlb
值越低,品种越抗
旱
(
方子云
, 2004,
长江科学院院报
, 21(4): 1-3;
张燕
之等
, 2002)
。因此
,
叶绿素含量的变化可作为衡量水
稻抗旱性强弱的重要指标
(
孙耀中和东方阳
, 2008)
。
转录因子
CBF
是一个的小基因家族
(Gilmour et
al., 1998)
。其
N
端含核定位信号区,
C
端含酸性激活
区,中间含与
DNA
结合的
AP2
结构域
(Haake et al.,
2002)
。
CBF
基因可分为两类,一类是依赖于
ABA
,
如
CBF4
;另一类是不依赖于
ABA
,如
CBF1
、
CBF2
、
CBF3
、
CBF5
、
CBF6(Sakuma et al., 2002)
。
CBF
基
因易受到低温、干旱、高盐等逆境的非生物胁迫因
子诱导表达
(Haake et al., 2002)
。在逆境胁迫下,
CBF