Basic HTML Version




分子植物育种
(
网络版
), 2012
年
,
第
10
卷
,
第
1164
-
1170
页
Fenzi Zhiwu Yuzhong (Online), 2012, Vol.10, 1164
-
1170
http://mpb.5th.sophiapublisher.com
1166
亮谷启动子区核苷酸多态性分布与前人报道的明
显不同。核苷酸多态性变异介于两组
(
类型
1,
类型
2;
类型
3,
类型
4,
类型
5,
类型
6,
类型
7)
之间,与
7
种类型的任何一种都不相似,而且发现
5
个特有
的
SNP
位点,分别位于-
1 851 bp
,-
1 720 bp
,-
700 bp
,
-
681 bp
和-
1 607 bp
处;
1
个特有的
InDel
,位于
-
523 bp
处。在启动子区的一些顺式作用元件
(cis-elements)
如
CCAAT
盒以及功能域如
ARR
没有
核苷酸变异,这与文献报道一致。
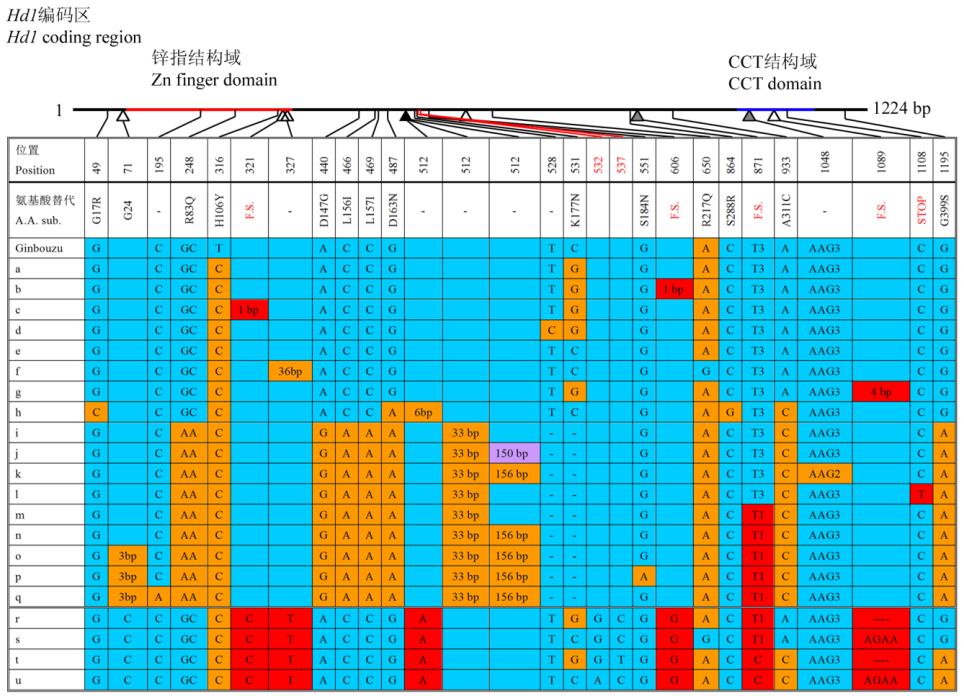
1.2 Hd1
编码区的核酸多态性
Hd1
编码区的核酸多态性分析结果显示
(
图
2)
,
Hd1
其具有很高的多态性。发现
6
个特有的
SNP/InDel
位点,其中包括了所有
4
个移码位点,
1
个插入位点和
1
个缺失位点。月亮谷
Hd1
基因包含
有前人报道的
871
位基因位点上的移码突变,从而
导致
Hd1
基因
CCT
结构域的部分失活,属无功能
的等位基因类型。而对前人报道的非移码突变及提
前终止位点,月亮谷
Hd1
基因类型介于上述类型之
间;
9311
属于无功能的类型
7
,日本晴则属于有功
能的类型
5
。
1.3
Ehd1
编码区的核酸多态性
Ehd1
编码区的核酸多态性分析的结果显示
(
图
3)
,月亮谷
Ehd1
编码区没有特有的
SNP/InDel
位点,与前人研究结果相似,可以归为类型
3
、类
型
4
、类型
5
、类型
6
和类型
7
组成的组。
1.4
Ghd7
等位基因的蛋白序列多态性
Ghd7
是影响每穗粒数、抽穗期的一个数量性
图
2
Hd1
核心编码序列区的多态性
注
: a-q:
类型
1
至类型
17; r: 9311; s:
日本晴
; t:
月亮谷-
1; u:
月亮谷-
2;
缺失和插入位点分别用空心和实心的三角箭头指示
;
F.S
表示阅读框架移码
;
核苷酸多态性用不同的颜色表示
; 9311
前数据引自
Takahashia
等
(2009)
Figure 2 A high polymorphism in the
Hd1
coding sequence
Note: a-q: Type 1 to type 17; r: 9311; s: Nipponbare; t: Yuelianggu
-
1; u: Yuelianggu
-
2; Deletion and insertion sites are shown by
open and closed arrowheads, respectively; F.S. is frame shift; Polymorphic nucleotides are indicated by different colors; The data
before 9311 is quoteg from Takahashiaet al. (2009)