Basic HTML Version





分子植物育种
(
网络版
), 2012
年
,
第
10
卷
,
第
1104
-
1114
页
Fenzi Zhiwu Yuzhong (Online), 2012, Vol.10, 1104
-
1114
http://mpb.5th.sophiapublisher.com
1105
2010)
。丝状病原菌在寄主植物内寄生,并能引发寄
主细胞坏死。病原菌与寄主植物间的互作分为亲和
性互作与非亲和性互作
(Liu et al., 2010)
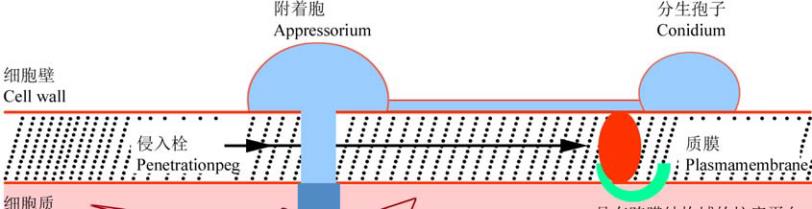
。根据病原
菌的效应子蛋白与寄主植物
R
蛋白的非亲和性作用
的位置,可将植物病原真菌分为
2
种类型:一种是
具有吸器的真菌,如大麦白粉菌与亚麻锈菌等;该
类病原菌的效应子蛋白可能先在其吸器中表达,然
后被病原菌转运到寄主的细胞质中;据推测其相应
的
R
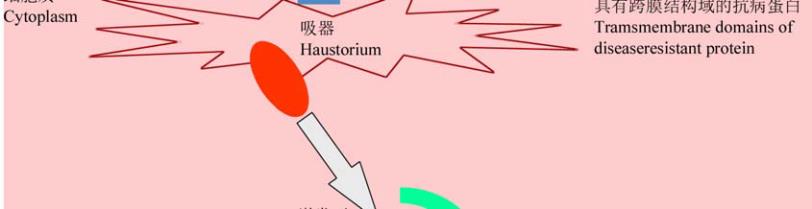
基因多位于细胞质。另一种是胞外分泌型真菌
(extracellular fungi)
,如番茄叶霉菌、油菜茎基溃疡
病菌、稻瘟病菌及大麦云纹病菌等;该类病原菌的
无毒基因被分泌到质外体,其相应的
R
基因多位于
细胞质膜上或具有一个跨膜结构域及一个胞间
LRR
结构
(
图
1) (Hammond-Kosack and Kanyuka, 2007;
de Wit et al., 2009)
。胞外分泌型病原菌最显著的特
征是编码胞外分泌型效应子蛋白。胞外分泌型效应
子具有如下特征;第一,该效应子被分泌到寄主植
物的质外体或木质部;第二,该效应子蛋白的
N
-
端,有时
C
-端往往在侵染期间被植物和
/
或真菌的
蛋白酶进一步地加工,形成成熟的蛋白;另外,同
其它在寄主细胞内活跃的效应子一样,胞外分泌型
效应子的另一个特征是,携有多个半胱氨酸残基,该
半胱氨酸残基可能参与维持蛋白结构稳定的分子内
二硫键的形成
(Stergiopoulos and de Wit, 2009)
。
由植物病原真菌引起的植物病害是农业生产
的主要限制因子之一,严重威胁着世界的农作物生
产。实践证明,培育和合理利用抗病品种是目前控
制此类病害最经济有效和对环境安全的手段。然
而,由于频繁出现能克服新导入的
R
基因的小种,
使得培育的抗病品种只具有短期的抗病效应
(Zeigler et al., 1994)
。随着生物信息学的快速发展以
及生物体全基因组测序时代的到来,这必将加速病
原菌无毒基因与寄主抗病基因的分离、克隆以及它
们之间相互作用分子机制的研究;从而为植物病害
的防治提供新的理论与途径。
本文综述了胞外分泌型真菌,包括番茄叶霉
菌、油菜茎基溃疡病菌及稻瘟病菌无毒基因研究的
最新进展。进一步对植物病原真菌无毒基因的结
构、内在
(intrinsic)
功能与进化机制以及后基因组时
代无毒基因克隆策略的变化及其对策进行了探讨。
图
1 Avr
蛋白和
R
蛋白在不同寄主—病原物互作过程中的位置
Figure 1 The location of Avr proteins and R proteins in different host-pathogen interaction