Basic HTML Version


刘淑君等
, 2011,
植物
MYB
转录因子的研究进展
,
分子植物育种
(online) Vol.9 No.114 pp.1835-1842 (doi: 10.5376/mpb. cn.2011.09.0114)
1836
物生长中的信号转导、次级代谢的调控、生物和非
生物胁迫的应答以及细胞周期的调控等等。由此可
见,
MYB
转录因子在植物的生长发育中扮演了重
要角色。
1 MYB
转录因子结构和分类
1.1 MYB
转录因子的结构特征
MYB
转录因子是指含有
MYB
基序的一类转录
因子,
MYB
基序约含
51
或
52
个氨基酸残基,其
中有三个保守的色氨酸残基,它们被
18
或
19
个氨
基酸残基序列所间隔。通常
MYB
基序形成三段螺
旋结构,且在第二段和第三段螺旋之间折转成一定
的 角 度 相 连 接 , 形 成 了 螺 旋
-
转 角
-
螺 旋
(helix-turn-helix, HTH)
结构
(Dubos et al., 2010)
,构
成了一个疏水核心,色氨酸对这个
HTH
结构有着
特别重要的意义。目前,第一个螺旋还没有明确的
结构意义,而第二个螺旋同
DNA
的磷酸骨架相接触,
稳固了与
DNA
的结合,第三个螺旋为识别螺旋,通
过氨基酸侧链以及
DNA
碱基之间的氢键同
DNA
序
列相识别
(Ogata et al., 1996;
王希庆等
, 2003)
。
原始的
MYB
基序通过自身的复制,产生了含
有
2
或
3
个
MYB
基序的
MYB
转录因子。为了更
好的协调与
DNA
结合的准确性,每个
MYB
基序
折叠成
HTH
形式,与
DNA
的大沟结合,而多个重
复的
MYB
基序会在与
DNA
结合处相互靠近,紧
密连接
(Weaver, 1999)
。
1.2 MYB
转录因子的分类
1987
年,
Martin
等人在玉米的糊粉粒中发现第
一个植物
MYB
转录因子
—
c-myb,
它参与花青素的
合成
(Marhamati et al., 1997)
。而后研究表明,
MYB
转录因子普遍存在于植物中,且在转录调节中起着
重要作用。以
c-MYB
为标准,将
MYB
基序分为
R1
、
R2
、
R3
三类
(Haga et al., 2007; Ogata et al., 1995;
Jia et al., 2004)
;另外,依据
MYB
基序重复种类和
数目的不同,将整个
MYB
转录因子家族分为四类
(Dubos et al., 2010)
—
4R-MYB
、
3R-MYB
、
1R-MYB/MYB-related
、
R2R3-MYB (
如图
1)
。
第一类是
4R-MYB
,由
4
个
R1/R2
重复组成,
它是植物
MYB
转录因子中最小的一类,仅在拟南
芥、杨树、葡萄
(Ito, 2005; Wilkins et al., 2009; Matus
et al., 2008)
中有所发现,且编码的蛋白质很少。
第二类是
3R-MYB (R1R2R3-MYB)
,由
R1
、
R2
、
R3
组成,普遍存在于大多数的真核生物中。
烟草中编码的
NtmybA1
、
NtmybA2
、
NtmybB
、
NtmybC1
和
NtmybC2
五个基因均属于此类转录因
子
(Jiang et al., 2004)
。
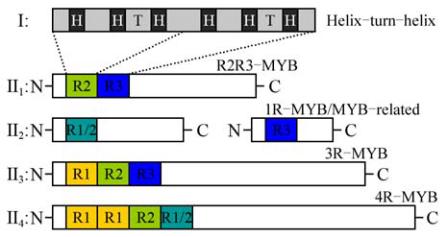
图
1
植物
MYB
转录因子的分类
(Dubos et al., 2010)
注
: I
:
MYB
基序的二级结构
;
螺旋
-
转角
-
螺旋
;
螺旋;转
角;
II:
四类
MYB
转录因子
: II1: R2R3-MYB; II2:
1R-MYB/MYB-related; II3: 3R-MYB, II4: 4R-MYB
Figure 1 Plant MYB transcription factor classes (Dubos et al.,
2010)
Note: I: MYB secondary structure: helix-turn-helix; H: helix; T:
turn; II: Four classes MYB transcription factor: II1:
R2R3-MYB; II2: 1R-MYB/MYB-related; II3: 3R-MYB, II4:
4R-MYB
第三类是
1R-MYB/MYB-related
,指含有单个
重复的或不完整的
MYB
基序,此类又可以分为
R1/R2-MYB
、
R3-MYB
、
MYB-related
三类。拟南
芥中
CCA1
和
LHY
属于
R1/R2-myb
类转录因子
,
由两个基因编码,参与生物钟的调节、光合作用基
因的表达以及激素的积累
(Carre et al., 2002)
;而后
鉴定的
CPC
属于
R3-MYB
类转录因子,与根毛的
形成有关
(Ohi et al., 1998)
。此外,研究发现
R3-MYB
很有可能是由于
R2R3-MYB
类转录因子中
R2
的缺
失而形成的
(Lipsick, 1996)
,此类转录因子大多数与
细胞形态的建成以及次级代谢的调节有关。
第四类是
R2R3-MYB
,为植物
MYB
转录因子
中最大的一类,几乎存在于所有的植物中。在模式
植物拟南芥中,
R2R3-MYB
类转录因子数目约占整
个
MYB
家族的
64.3% (
刘翔等
, 2010)
。
R2R3-MYB
类转录因子不仅数目众多,而且功能具有多样化。
目前,在
R2R3-MYB
类转录因子中,
N
末端为
DNA
结合区域,
C
末端为转录调控区域
(Dubos et al.,
2010)
。
DNA
结合区域的氨基酸序列比较保守,决
定了转录因子与
DNA
结合的特异性;转录调控区
域包括转录激活区和转录抑制区,而有的转录因子