Basic HTML Version




水生生物研究
, 2012
年
,
第
1
卷
,
第
7-13
页
Shuisheng Shengwu Yanjiu, 2012, Vol.1, 7-13
http://aor.5th.sophiapublisher.com
9
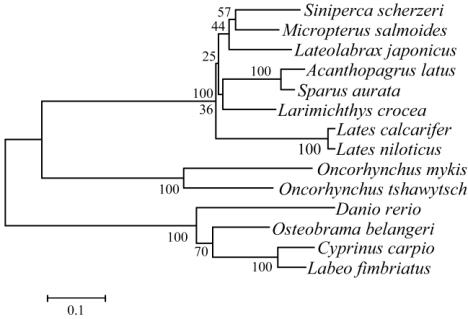
目鲑科的虹鳟鱼、大鳞大麻哈鱼分别在另两个大
的分支上。图中
myostatin
启动子进化树所表明的
鱼类亲缘关系及其本身的分类地位,与传统形态
学分类结果相一致。
图
3 14
种鱼类
myostatin
启动子序列的
NJ
系统进化树
注
:
大口黑鲈
: EF071854;
斑马鱼
: AY323521;
虹鳟鱼
:
DQ138300;
大黄鱼
: AY842934;
花鲈
: AY965685;
鲤鱼
:
GU014395;
缨野鲮
: HQ850576;
尖吻鲈
: EF672685;
尼罗尖
吻鲈
: EF681885;
金头鲷
: EU881511;
黄鳍棘鲷
: HM368446;
比氏骨鳊
: HQ850577;
大鳞大麻哈鱼
: EU009954
Figure 3 The NJ phylogenetic tree of the myostatin promoter
sequences from 14 kinds of fishes
Note:
Micropterus salmoides
: EF071854;
Danio rerio
:
AY323521;
Oncorhynchus mykiss
: DQ138300;
Larimichthys
croce
a: AY842934;
Lateolabrax japonicus
: AY965685;
Cyprinus carpio
: GU014395;
Labeo fimbriatus
: HQ850576;
Lates calcarifer
: EF672685;
Lates niloticus
: EF681885;
Sparus
aurata
: EU881511;
Osteobrama belangeri
: HQ850577;
Acanthopagrus latus
: HM368446;
Oncorhynchus tshawytscha
:
EU009954
2
讨论
Tail-PCR
是一种用来克隆已知序列两端的未知序列
的新方法,特异性高,重复性好,能快速获得目标
片段。避免了基因组步行法对高质量基因组
DNA
的要求,与普通
PCR
一样,可以使用低质量的基因
组
DNA
做模板。本试验利用
Tail-PCR
的方法克隆
出斑鳜
myostatin
的上游调控序列,长度约为
860
bp
。
PLACE
预测结果显示,该基因上游调控区除含
有一个核心启动序列
(732~781 bp)
外,还有
I-box
(156~160 bp)
、
TATAA box (730~744 bp)
、
CAAT-box
(698~701 bp)
及
4
个
E-box (26~31 bp, 287~292 bp,
589~594 bp, 608~613 bp)
等启动子特征性调控元件
的存在。进一步分析表明:斑鳜
E-box3
是
MyoD
转录因子结合位点,
MyoD
可通过与
E-box
结合调
控其下游靶基因
myostatin
的转录和表达,对肌肉
的生长发育有着重要的调控作用
(Langley et al.,
2002; Østbye et al., 2007)
。作为
MyoD
家族转录因
子结合位点的
E-box
元件同样在斑马鱼
(Xu et al.,
2003)
、石斑鱼
(Chen et al., 2007)
、虹鳟
(Garikipati et
al., 2006)
、花鲈
(Ye et al., 2007)
、大西洋鲑
(Østbye et
al., 2007)
等硬骨鱼的
myostatin
启动子序列中被发
现。调控序列中的
CAAT-box
元件可极大提高基本
启动子的转录效率
(
孙晓红等
, 2002)
,对
myostatin
基因的转录水平也有着较大的影响。
对
myostatin
启动子的进化分析表明:斑鳜
myostatin
启动子与其它
13
种鱼亲缘关系的远近和
本身的分类学地位,与传统形态学分类结果相一
致。由此我们可以假设,启动子的变化与物种的进
化有着相关性,启动子通过碱基变化来改变结构基
因的转录水平从而促使物种向着与启动子作用相
关的方向进化。
DNA
序列分析表明,克隆得到的斑鳜
myostatin
结构基因长
2 360 bp
,由
3
个外显子和
2
个
内含子组成,外显子大小依次为
379 bp
、
371 bp
和
381 bp
;内含
I
和内含子
II
大小分别为
395 bp
和
830 bp
。两个内含子都以
GT
开始、
AG
结尾,符合
内含子的
GT-AG
剪接规则。进一步分析发现:内含
子
I
序列中有
2
个
CAAT-box
,内含子
II
序列中有
1
个
TATAA-box
,
5
个
CAAT-box
等类似启动子调控元件,
推测斑鳜
myostatin
基因内含子可能具有增强该基
因转录的功能。同样,典型的调控元件
CArG
和
TATAA-box
序列也分别在鲮
β-actin
基因内含子
1
和
准噶尔雅罗鱼
β-actin
基因的第
1
个内含子中被发现,
预示着这些内含子可能作为相应基因启动子的一
部分在起始转录表达中有着重要作用
(
张殿昌等
,
2007;
胡文革等
, 2010)
。
斑鳜
myostatin
基因推导的氨基酸序列显示:
该蛋白在
22~23
位氨基酸之间存在一个信号肽切割
位点,
264~267
位氨基酸处有
RARR (
与
RXXR
相
一致
)
蛋白酶解的加工位点。与其它
TGF-β
超家族
成员蛋白一样,
myostatin
蛋白以前体形式存在,切
除信号肽后,在
RXXR
位点经蛋白酶水解加工,形
成
N
端编码前肽和
C
端生物活性区
(Lee and
McPherron, 2001)
。斑鳜
myostatin
氨基酸序列
C
端
生物活性区的保守性很高,其中有
9
个异常保守的
半胱氨酸,它们以二硫键形式连接形成的
“cysteine
knot”
结构可能与
myostatin
蛋白及其它
TGF-β
超家