Basic HTML Version


郑燕等
, 2011,
一个水稻长穗大粒特异种质的主效
QTL
检测和定位
,
分子植物育种
Vol.9 No.13 (doi: 10.5376/mpb.cn.2011.09.0013)
1096
所有的
QTL
都定位出来。但在育种实践中,要同时
对许多
QTL
进行选择是十分困难的,缺乏可操作
性。好在已有的研究表明,每个数量性状通常都存
在几个主效
QTL
,它们往往能够解释大部分的遗传
变异。因此,从育种角度出发,检测和定位主效
QTL
更有实际意义。而从理论上看,主效
QTL
不仅更容
易研究,而且可能更有助于对目标性状分子遗传机
理的理解。目前已有
2
个控制粒长的主效
QTL (
LK
-
4
和
gl
-
3
)
和
2
个控制粒重的主效
QTL (
gw3.1
和
gw3.8
)
被精细定位
(Zhou et al., 2006; Wang et al., 2006; Li
et al., 2004; Xie et al., 2006)
;有
2
个控制粒长和粒重
的主效
QTL (
GS3
和
GW2
)
被克隆,它们分别编码一
种跨膜蛋白和
RING-type E3
泛素连接酶
(Fan et al.,
2006; Song et al., 2007)
。
在遗传研究中,特殊的种质往往具有重要的价
值。选择极端类型的材料作为杂交亲本,更有利于
发现主效
QTL
。我们自
1995
年起引进国际水稻所新
株型水稻品系,通过杂交、复交和辐射等育种手段,
获得一批长穗大粒特异种质。这些种质对研究水稻
穗粒形态性状的遗传基础无疑是十分有用的。本研
究对一个长穗大粒特异种质
LPBG08
中控制穗长、
粒长和粒宽的主效
QTL
进行检测和定位,以期为水
稻穗粒性状的分子标记辅助育种和分子遗传研究
奠定基础。
1
结果与分析
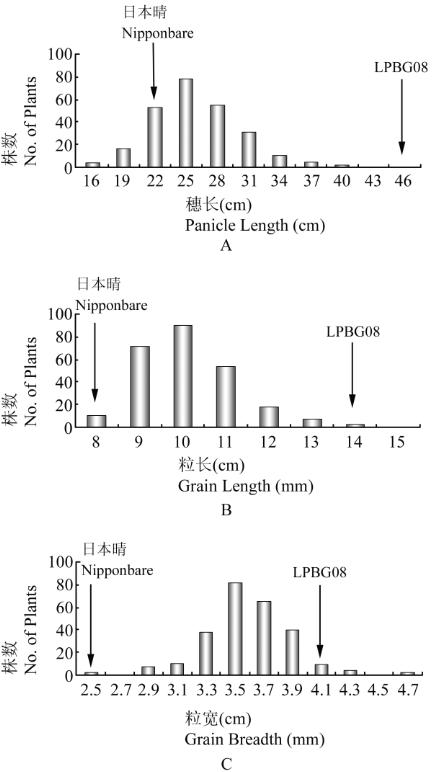
亲本
LPBG08
的穗长接近
46 cm
,粒长达
15.0 mm
,
粒宽达
4.1 mm
,是一个典型的长穗大粒的品系,与
另一个亲本日本晴之间存在很大的差异
(
图
1)
。
F
2
中
3
个性状皆出现广泛的分离,并存在超亲遗传的现
象,但都表现为连续变异,接近正态分布
(
图
1)
。说
明尽管两亲本间差异很大,
3
个性状仍表现为典型
的数量遗传。
用
1 039
对
SSR
引物对两亲本进行检测,共筛选
到
265
对多态引物,比例为
25.5%
。用多态引物对各
对
DNA
池进行检测,结果位于
1
号染色体上的
RM572
在宽粒池与窄粒池之间表现多态;位于
3
号
染色体上的
RM448
在长穗池与短穗池之间及长粒
池与短粒池之间均表现多态。故推测在这
2
个标记
附近分别存在控制粒宽和穗长及粒长的
QTL
。于是
基于所构建的局域连锁图,共定位了
4
个
QTL(
图
2,
图
1 F
2
群体中穗长、粒长和粒宽的频率分布
注
:
箭头所指为亲本的均值
; A:
穗长
; B:
粒长
; C:
粒宽
Figure 1 Frequency distributions of panicle length, grain length
and grain breadth in the F
2
population
Note: Arrows indicate the mean values of the parents; A:
Panicle length; B: Grain length; C: Grain breadth
表
1)
,其中在
1
号染色体上定位到
1
个控制粒宽的
QTL (
qGB1
)
,在
3
号染色体上分别定位到
1
个控制粒
长
QTL (
qGL3
)
和
2
个控制穗长的
QTL (
qPL3
-
1
和
qPL3
-
2
)
。所有
QTL
的增效基因都来自于
LPBG08
,
且加性效应对表型变异的贡献率都超过
10%
,其中
qGL3
和
qPL3
-
1
的贡献率较大。但
qGL3
和
qPL3
-
1
的
显性效应都不明显,说明这
2
个
QTL
主要表现为加
性作用模式。
qGB1
和
qPL3
-
2
则表现出明显的显性
效应,其中
qGB1
表现为完全显性
(
显性度为
1.13)
,
显性方向为增效;而
qPL3
-
2
表现为超显性
(
显性度为