Basic HTML Version




水生生物研究
, 2012
年
,
第
1
卷
,
第
1-6
页
Shuisheng Shengwu Yanjiu, 2012, Vol.1, 1-6
http://aor.5th.sophiapublisher.com
2
着养殖业的发展,中华鳖种群混杂导致种群间的分
化、变异逐渐被人为湮灭,其种群的遗传多样性受
到了很大的影响。刘至治等
(2004)
应用
RAPD
分析了
中华鳖五群体的遗传变异,认为遗传多样性较丰
富;肖亚梅等
(2005)
研究得出中华鳖群体内的遗传
变异率
0.227
,为遗传共享度较高的群体;张志允等
(2011)
对中华鳖黄河群体选育世代
F
1
、
F
2
及
F
3
遗传变
异进行了微卫星分析,结果表明群体内存在丰富可
供选育用的遗传多样性基础,随着选育代数的增加
而明显降低,证明以生长、体色和体型等表型指标
为直观选育指标的群体选育,对遗传型指标产生了
可检测到的影响。
遗传多样性大小是物种长期进化的产物,是其
生存适应和发展进化的前提。一个居群或物种遗传
多样性越高或遗传变异越丰富,对环境变化的适应
能力就越强。了解物种的遗传变异,有利于对其种
质资源的管理保护和开发利用。因此,本研究利用
微卫星标记对
5
个养殖种群中华鳖的遗传多样性及
种群间的分化进行分析,以期为中华鳖的种质资源
保护以及改良育种工作提供理论依据。
1
结果与分析
1.1
微卫星扩增结果
试验所用
11
对微卫星引物均能在所有
DNA
样
品中稳定地扩增出相应条带
(
图
1)
。各微卫星引物在
5
个群体
150
个个体中的等位基因数为
3~6
个,平
均等位基因数为
4.181 8
,平均有效等位基因数为
2.229 3
,扩增片段大小在
108~246
之间。洞庭
(DT)
、
黄河
(HH)
、黄沙
(HS)
、日本
(RB)
以及绿卡
(LK)
在各
个位点的等位基因数及有效等位基因数见表
2
。各
个群体的平均有效等位基因数、平均等位基因数、
平均观测杂合度、平均期望杂合度以及平均多态信
息含量如表
3
所示。
1.2
遗传多样性分析
11
个位点中
P
-
05
、
P
-
O6
、
P
-
12
、
P
-
11
、
P
-
12
和
PS
-
04
位点的多态信息含量
(PIC)
小于
0.5
,为中
度或者低度多态,其余位点均为高度多态,说明微
卫星引物多态性较好,可以用于遗传多样性分析。
根据每个位点的等位基因频率,计算反映群体遗传
多样性的有效等位基因数、观测杂合度、期望杂合
度以及多态信息含量,其平均数值范围分别为
2.018 9~
2.396 9
、
0.433 3~0.554 5
、
0.427 8~0.527 4
和
0.402 0~
0.466 0
。平均有效等位基因数、平均观测杂合度、
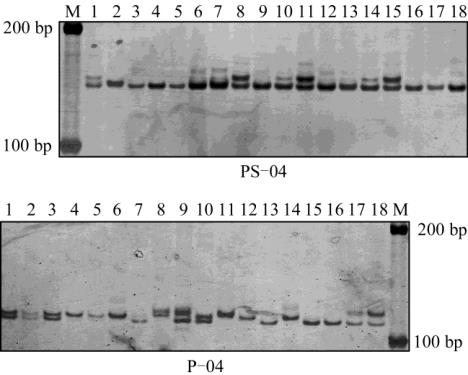
图
1
引物
P
-
04
和
PS
-
04
在中华鳖五个群体部分个体中的扩增
带谱
Figure 1 The example of band pattern amplified by primers of
P
-
04 and PS
-
04 in Trionyx sinensis
平均期望杂合度以及平均多态信息含量均为杂交
种群绿卡
(LK)
的最高,分别为
2.396 9
、
0.554 5
、
0.527
4
和
0.466 0
;而以日本
(RB)
种群的最低,分别为
2.019
8
、
0.433 3
、
0.427 8
和
0.402 0
。这些指标的分析结
果表明绿卡
(LK)
的遗传多样性最高,洞庭、黄河和
黄沙次之,日本
(RB)
的遗传多样性最低。
1.3
群体间遗传结构分析
通过对
5
个群体中华鳖进行
F
-检验
(
表
4)
,结
果显示洞庭
(DT)
和日本
(RB)
的遗传分化指数最大
(FST=0.096 9)
,而洞庭
(DT)
和黄河
(HH)
的分化指数
最小
(FST=0.023 3)
。遗传相似率和遗传距离如表
5
。
根据亲本与子代间的遗传距离,采用
UPGMA
法构
建子代与亲本的系统进化树
(
图
2)
,由图可知,黄河
(HH)
和洞庭
(DT)
先聚为一支,然后与其子代绿卡
(LK)
聚合,再与黄沙
(HS)
聚合,最后才与日本
(RB)
聚合。
2
讨论
2.1
中华鳖
5
个群体的遗传多样性
多态信息含量
(PIC)
是群体内遗传变异的量度,
可以用来描述微卫星位点的变异程度。依
Botstein
等的划分标准:当
PIC>0.5
时,该位点为高度多态
位点;当
0.25<PIC<0.5
时,为中度多态性位点;当
PIC<0.25
时,为低度多态位点。本研究的结果显示,
中华鳖
5
个种群的多态信息含量都处于中度水平,