Basic HTML Version


张广辉等
, 2011,
茶树儿茶素酯化分子机理与研究途径
,
分子植物育种
Vol.9 No.71 (doi: 10.5376/mpb.cn.2011.09.0071)
1519
酯型儿茶素,其余为非酯型儿茶素
(
简单儿茶素
)
。
1
酯型儿茶素是茶树的特有类黄酮物质
积累高含量的酯型儿茶素是茶树类黄酮生物
合成与代谢的最大特点。简单儿茶素普遍存在于植
物体内,是合成原花色素
(proanthocyanidin)
的前体
物质
(He et al., 2008; Aron and Kennedy, 2008)
,但酯
型儿茶素仅专一地、大量存在于山茶科山茶属茶组
(Section
Thea
)
植物,如茶树
[
Camellia sinensis
(L.) O.
Kuntze]
、普洱茶变种
[
C. sinensis
var.
assamica
(Masters) Kitamura]
和大理茶
(
C. taliensis
)
中,茶组的
滇缅茶
(
C. irrawadiensis
)
仅含少量酯型儿茶素,其他
山茶属植物仅含简单儿茶素
(Nagata and Sakai, 1984;
Yagi et al., 2009; Li et al., 2010)
。除茶树外,酯型儿
茶素仅在葡萄
(
Vitis vinifera
)
种子中
(Guendez et al.,
2005)
有少量存在。
茶树和普洱茶变种中,酯型儿茶素含量占茶多
酚总量的
48.7%~91.1% (Li et al., 2010)
,研究证实
EGCG
能够调节多种人类疾病特异目标分子
(Nagle
et al., 2006)
。酯型儿茶素仅存在于茶组少数植物中,
说明这些植物体内存在独特的儿茶素酯化反应,并
且催化该反应的酶也仅特异地存在这些植物体内。
2
儿茶素酯化机理可能的分子机理
植物类黄酮
(
包括黄烷-
3
-醇
)
的生物合成与调
节 机 理 已 经 比 较 清 楚
(Winkel-Shirley, 2001;
Forkmann and Martens, 2001)
。近年茶树类黄酮生物
合成研究集中于结构基因的克隆与功能分析
(Park
et al., 2004; Punyasiri et al., 2004; Lin et al., 2007;
Singh et al., 2008; 2009a; 2009b;
马春雷等
, 2010)
,
少量儿茶素积累与结构基因表达的关系研究
(Eungwanichayapant and Popluechai, 2009; Ashihara
et al., 2010)
。
茶树酯型儿茶素的酯化生物合成机理一直为
研究者所忽视,甚至催化其反应的酶也尚未命名。
仅见的研究表明,茶树嫩梢饲喂没食子酸-
G-C
14
后,
能够在
EGCG
和
ECG
中检测到放射性,说明酯型儿
茶素由没食子酸和简单儿茶素酯化而形成的
(Saijo,
1983)
。由于茶树体内没食子酸含量很低,而儿茶素
酯化需要大量没食子酸,说明儿茶素酯化反应进行
非常迅速
(Saijo, 1983; Ashihara et al., 2010)
,暗示编
码该酶的基因转录水平高或者酶的催化效率高。
Ashihara
等
(2010)
将催化儿茶素酯化的酶称为黄烷
-
3
醇没食子酸酯合成酶
(flavan
-
3
-
ol gallate synthase,
FGS)
,但该名称未表明其反应机理。参照催化其它
酯化反应的酶的命名,本文将催化儿茶素酯化的酶
暂定名为黄烷-
3
-醇没食子酰基转移酶
(flavan
-
3
-
ol
galloyltransferase, GaT)
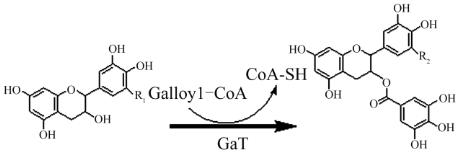
,其可能的反应如图
1
所示。
图
1
可能的儿茶素酯化反应机理
注
: GaT:
没食子酰-
CoA:
黄烷-
3
-醇没食子酰基转移酶
; EC:
R
1
=H, (-)
-表儿茶素
, EGC; R
1
=OH, (-)
-表没食子儿茶素
;
ECG: R
2
=H, (-)
-表儿茶素没食子酸酯
; EGCG: R
2
=OH, (-)
-
表没食子儿茶素没食子酸酯
Figure 1 The assumed reaction mechanism of catechins
esterfication
Note: GaT: galloyl-coenzyme A:flavan
-
3
-
ol galloyltransferase;
EC: R
1
=H, (-)-epicatechin, EGC; R
1
=OH, (-)-epigallocatechin;
ECG: R
2
=H, (-)-epicatechin gallate; EGCG: R
2
=OH,
(-)-epigallocatechin gallate
3
克隆茶树
GaT
基因可能的途径
如上所述酯型儿茶素仅存在于茶树等少数植
物体内,因此很难利用克隆同源基因的方法来克隆
茶树
GaT
基因。近年一些植物酰基转移酶基因克隆
成功,为克隆茶树
GaT
基因提供了借鉴。
根据儿茶素酯化反应的底物与产物分子结构,
可知催化该反应的酶属于植物酰基转移酶的
BAHD
家族。
BAHD
酰基转移酶家族利用
CoA
硫酯,催化
形成多种植物次生代谢物。
BAHD
家族包括
4
种酰基
转移酶,即
2
个乙酰基转移酶,分别是苯甲醇
O
-乙
酰基转移酶
(benzylalcohol
O
-acetyltransferase, BEAT)
和去乙酰基文多灵
4
-
O
-乙酰基转移酶
(deacetylvin-
doline 4
-
O
-
acetyltransferase, DAT),
前者催化合成花
香物质乙酸苯甲酯和生物碱文多灵
(vindoline)
。另
外
2
个是苯甲酰
/
羟基肉桂酰
CoA
酰基转移酶
(benzoyl/hydroxyl-cinnamoyl CoA acyltransferases)
,
分别是邻氨基苯甲酸
N
-羟基肉桂酰
/
苯甲酰基转移
酶
(anthranilate
N
-hydroxycinnamoyl/benzoyltransferase,
HCBT)
和花青素
O
-羟基肉桂酰转移酶
(anthocyanin