Basic HTML Version


束红梅等
, 2011,
油菜素内酯代谢相关基因及其调控植物耐盐性的研究进展
,
分子植物育种
Vol.9 No.34 (doi: 10.5376/mpb.cn.2011.09.0034)
1262
et al., 2001;
曹云英等
, 2007;
万正林等
, 2009)
。研究
表明,在盐胁迫条件下外施油菜素内酯可提高植物
的耐盐性
(Bajguz and Hayat, 2009)
,但是相关的耐盐
机理尚未明确。
油菜素内酯除了在植株地上部合成外,在根系
同样可以合成,其在植物体内没有长距离的运输,
主要在合成部位原位起作用
(Sakurai, 1999; Symons
and Reid, 2004; Bajguz, 2007)
。而且油菜素内酯的合
成代谢过程涉及多个步骤,且每个步骤均受到相关
基因的调控。因此,本文综述了油菜素内酯的生物
代谢途径及相关基因功能及其在提高植物耐盐性
方面的研究现状,并对油菜素内酯提高植物耐盐机
理研究工作的开展进行展望。
1
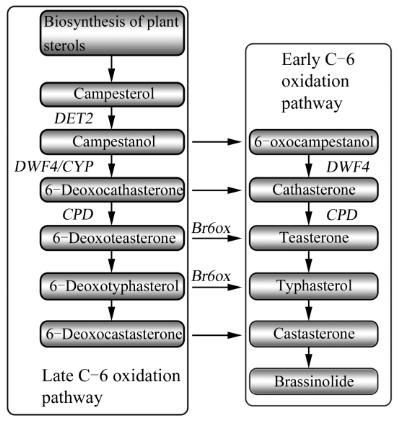
油菜素内酯的合成途径
研究表明,菜油甾醇
(Campesterol)
是油菜素内
酯生物合成的起始物,在
DET2
基因催化后得到菜油
甾烷醇
(Campestanol)
,菜油甾烷醇在甾醇体和侧链
上发生一系列羟化和氧化步骤的同时伴随着
C
-
6
位
置的酮基化,这种酮基化发生在
C
-
22
、
C
-
23
、
C
-
3
和
C
-
2
位置的修饰前和后
(Bishop and Yokota, 2001;
Shimada et al., 2001;
储昭庆
, 2006)
,这两条途径分
别被称为早期
C
-
6
氧化途径和晚期
C
-
6
氧化途径
(Noguchi et al., 2000; Bishop and Koncz, 2002; Kim
et al., 2005;
储昭庆
, 2006) (
合成途径见图
1, Divi and
Krishna, 2009)
。
1.1
早期
C6
氧化途径
菜油甾醇
(Campesterol)
作为油菜素内酯生物合
成的起始物,经加氧、
6α
-羟化、氧化等得到
6
-氧
菜油甾烷醇
(6
-
oxocampestanol)
,再经羟化得茶甾酮
(Teasterone)
,经脱羟基、再羟化为香蒲甾醇
(Typhasterol)
,接着转化为油菜素甾酮
(Castasterone)
、
油菜素内酯,这个合成途径称之为早期
C6
氧化途径
(
储昭庆等
, 2006;
罗明
, 2007; Divi and Krishna,
2009)
。拟南芥、烟草等多种植物中均存在这一油菜
素内酯合成途径
(Noguchi et al., 2000)
,说明油菜素
内酯的早期
C
-
6
氧化途径在植物中广泛存在。
1.2
后期
C6
氧化途径
以前人们因为
6
-脱氧油菜素内酯的生物活性
较低,认为其不能转化为活性油菜素内酯。但近年
来的研究发现在多种植物中
6
-脱氧油菜素内酯均
图
1
油菜素内酯合成过程简图
Figure 1 The brassinosteriod biosynthetic pathway
参与了油菜素内酯的生物合成过程
(Choi et al., 1997;
罗明
, 2007)
,并将这一生物合成途径称之为后期
C
-
6
氧化途径。该途径同样以菜油甾醇
(Campesterol)
为起始物,经氧化等到菜油甾烷醇
(Campestanol)
,
再经羟化得
6
-脱氧长春花甾酮
(6
-
Deoxocathasterone)
、
6
-脱氧茶甾酮
(6
-
Deoxoteasterone)
,经脱羟基、再羟
化为
6
-脱氧香蒲甾醇
(6
-
Deoxotyphasterol)
,接着转
化为
6
-脱氧油菜素甾酮
(6
-
Deoxocastasterone)
、油菜
素甾酮、油菜素内酯
(Choi et al., 1997; Jager et al.,
2007)
,而且这一途径也存在于许多植物中。
2
油菜素内酯合成相关基因
从油菜素内酯的合成途径可以看出该过程涉
及到多个步骤,而这些步骤受到多个基因的调控,
下面主要概述油菜素内酯合成过程中一些主要基
因及其功能。
2.1
DET2
DET2
基因编码油菜素内酯生物合成酶,研究发
现其编码的蛋白与哺乳动物中类固醇
5α
-还原酶的
同源性极高
(Li et al., 1996)
。通过研究发现,突变体
不能把同位素标记的菜油甾醇转化为菜油甾烷醇,
而位于
DET2
催化反应步骤之后的所有油菜素内酯
合成途径中的中间产物都可以逆转
det2
的突变表型
(Asami and Yoshida, 1999)
。说明
DET2
催化菜油甾醇
转化为菜油甾烷醇,是油菜素内酯合成过程中催化