Basic HTML Version



Journal of Mosquito Research, 2013, Vol.3, No.5, 33
-
44
http://jmr.sophiapublisher.com
35
insect larvae were washed from nematode and
transferred to new Petri dishes and were incubated for
further 72 hr. The Percentage of larval mortalities was
determined and the number of nematodes present in
each cadaver was determined by dissection after 3,6,9
and 12 hr according to Mauleon et al.(1993).
2.6 Dose response assay
In the present assay, the effect of different concentr-
ations of infective juvenile nematodes of
H. bacteri-
ophora
and
H. indica
on the mortality of 4
th
instar
mosquito larvae was monitored and LC
50
values were
calculated for both nematode species. The nematode
concentrations used include 0 (control),100,150,200,
250 and 300 ij/larva. The insect mortality was recorded
48 hr post-inoculation.
To study the effect of host larval instar on infection by
nematode and hence the resulted host mortality, the
experiment was repeated using younger host instar
larvae (2
nd
, 3
rd
) using the infective juveniles of
H.
bacteriophora
.
2.7 Nematode fecundity
Infective juvenile production was used as an indirect
measure to first-generation nematode fecundity. The
used nematode concentrations were (50,100,150,200,
250 and 300 ij/larva). Host cadavers (three replicates)
were transferred to individual White dishes (Woodring
& Kaya, 1988). All infective juveniles that emerged
from a single host at 48, 72 and 96 hr post exposure
were collected and counted.
The first generation of
H. bacteriophora
and
H. indica
within the host is represented by hermaphrodites.
Because body length is correlated with fecundity
(Selvan et al., 1993), the total length of 12 hermaphro-
dites at variable infective juvenile concentrations (50,
100,150,200,250 and 300 ij/larva) was measured at 48,
72 and 96 hr post exposure. The control value (0 nem-
atode in host) is based on hermaphrodite length that
emerged from the rearing host,
Galleria mellonella
larvae. Since both species showed similar recovery
pattern, only
H. bacteriophora
was selected in this
assay. Infective juvenile length was also measured for
each concentration. Prior to measurements, nematodes
were heat killed on a glass slide.
2.8 Statistical analyses
In bioassay studies, host larval mortalities were calcul-
ated according to Abbott (1925). Values of LC
50
and
LT 50 were calculated according to Finney (1971)
using Probit analysis (SPSS, v11). Hermaphrodite
length and female fecundity were analyzed by general
linear models procedures. There were five replicates
per treatment.
All comparisons were made at the 0.05 level of signif-
icance. Data were presented as means ± standard error.
Comparisons by means were done by student t-test
and one-way analysis of variance.
3 Results and Analysis
3.1 Susceptibility to EPN
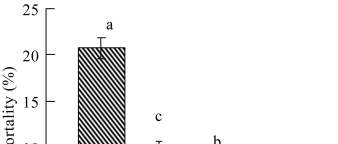
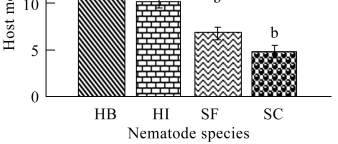
A significant increase in host larval mortality was ach-
ieved by using both
Heterorhabditis
species, where
H.
bacteriophora
was significantly more virulent than
H.
indica
(P
≤
0.05) (Figure 1). Both
Heterorhabditis
species
raised the host mortality levels above the normal
control values by a magnitude of 2~4 times. In contrast,
the mortality levels did not exceed 7% in case of using
both
Steinernema
strains, with no significant differentce
in host mortality between both strains (P
≥
0.05). The
host mortality levels achieved by applying both
Stein-
ernema sp
. were nearly doubled when
H. indica
was
used. In the same time,
H. bacteriophora
was proved to
be the most virulent species, where the recorded host
mortality levels were 2 times the mortality values ach-
Figure 1 The percentage mortality of
C. quinquefasciatus
larvae following exposure for 72 hr to individual ijs of
H.
bacteriophora
(=HB),
H. indica
(=HI),
S. carpocapsae
(=SC)
and
S. feltiae
(=SF) in the one-on-one assay. Means with the
same letter are not significantly different (P
≥
0.05)