Basic HTML Version



Plant Gene and Trait, 2013, Vol.4, No.20, 109
-
123
http://pgt.sophiapublisher.com
110
Singh et al., 2011). Upon exposure to water stress,
plants dramatically increase the production of
reactive oxygen species (ROS) such as singlet oxygen
(
1
O
2
), superoxide (O
2
•−
), hydrogen peroxide (H
2
O
2
),
and the hydroxyl radical (•OH) in different subcellular
organelles (Cruz de Carvalho, 2008). Restricted
entries of CO
2
in the leaves during drought stress limit
the CO
2
fixation and accelerate the photorespiratory
pathway and finally lead to excessive H
2
O
2
in the
peroxisome (Cruz de Carvalho, 2008). It has been
estimated that under drought stress more than 70% of
total H
2
O
2
accumulation is due to photorespiration
(Noctor et al., 2002). Although ROS mainly H
2
O
2
can
act as signals to help plants adapt to stress responses
(Miller et al., 2008, 2010; Gill and Tuteja, 2010;
Hossain and Fujita, 2012; Petrov and Van Breusegem,
2012), excess ROS cause oxidative damages to plant
macromolecules (Gill and Tuteja, 2010; Jaspers and
Kangasjarvi, 2010; Hossain et al., 2011a). As a result
metabolic alteration, inhibition of photosynthesis,
and breakdown of cellular organization contribute to
growth retardation, reduced fertility, premature
senescence even death of plants (Saito et al., 2010;
Krasensky and Jonak, 2010; Hossain et al., 2011a).
Therefore, the level of ROS should be judiciously
regulated in plants through the coordinate of ROS
production system and ROS scavenging system to
manage oxidative damage and simultaneously
regulate signalling events (Foyer and Noctor, 2005;
Rouhier et al., 2008), but the fundamental
mechanisms are still largely unknown.
Methylglyoxal (MG) and abiotic stresses are
inextricably intertwined, the over-accumulation and
higher rate of MG production in plants possibly being
a general stress response (Hossain et al., 2009; Banu
et al., 2010; Upadhyaya et al., 2011). MG is a typical
α-oxoaldehyde, which forms as a by-product of
several metabolic pathways, e.g. glycolysis, lipid
peroxidation and oxidative degradation of glucose
and glycated proteins. It is toxic to plant cells,
causing inhibition of cell proliferation, degradation
of proteins and inactivation of antioxidant defense
systems and as a consequence disrupts cellular
functions (Matins et al., 2001; Hoque et al., 2010,
2012a). MG accumulates in plants under normal plant
physiological processes like photosynthesis and MG
vividly elevated under various stress conditions
(Yadav et al. 2005a; Hossain et al., 2009). Recently
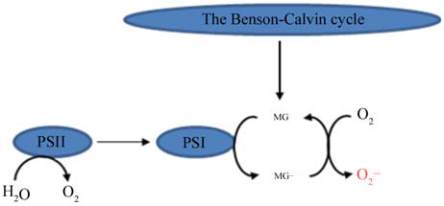
Saito et al. (2011) reported that MG accumulated in
chloroplasts during the day from triose phoshphates,
needs to be controlled by detoxification mechanisms,
otherwise it will catalyse the photoreduction of O
2
to
O
2
•−
at photosystem I (Figure 1) and the increase in
O
2
•−
production during photosynthesis further
aggravate the oxidative damage to plant cells. MG
not only directly inhibits physiological functioning
but it also inhibits it via the changes in ABA
synthesis in
Arabidopsis
(Hoque et al., 2012b,
2012c). Therefore, in order to survive under
stressful conditions plants must up-regulate MG
detoxification process to avoid cellular damage and
also to keep steady state pace in different plant
physiological processes.
Figure 1 Methylglyoxal induced enhancement of superoxide
production (O
2 •−
) in chloroplast (modified from Saito et al.,
2010)
To avoid the excessive accumulation of ROS and to
protect from oxidative damage, plants are armed with
sophisticated antioxidant defense systems: both
enzymatic antioxidant (multiple superoxide
dismutases (SOD), ascorbate peroxidase (APX),
monodehydroascorbate reductase (MDHAR),
dehydroascorbate reductase (DHAR), glutathione
reductase (GR), catalase (CAT), glutathione
peroxidase (GPX), glutathione
S
-transferase (GST),
peroxidase (POX), guaiacol peroxidase (GPOX) and
non-enzymatic
antioxidant
(asocrbare
(AsA),
glutathione (GSH), tocopherol, carotenoids, flavonoids,
and proline)(Figure 2; Gill and Tuteja, 2010; Hossain
et al., 2011a; Sharma et al., 2012) in different
sub-cellular organelles. The delicate balance
between ROS production and scavenging that
allows this duality in function to exist in plants is
thought to be orchestrated by a large network of
genes that tightly regulates ROS production and
scavenging (Mittler et al., 2004; Miller et al., 2008;
Petrov and Van Breusegem, 2012).