Basic HTML Version



International Journal of Marine Science 2013, Vol.3, No.9, 72-78
http://ijms.sophiapublisher.com
73
sequestration, as anaerobic conditions prevailing in
the soils due to frequent flooding reduces its oxidation
and limits release into the atmosphere as CO
2
.
Total of 20 true mangrove species, belong to 11
plant families were identified in Sri Lanka (Jayatissa
et al., 2002). Rhizophoraceace, Combretaceae,
Euphorbiaceae and Avicenniaceae are abundant in Sri
Lanka.
Bruguiera gymnorrhiza,
which belong to the
family Rhizophoraceace and is one of the most
distributed pantropical families in the world
(Tomlinson, 1986), and
Lumnitzera racemosa,
which
belongs to family Combretaceae, were selected for the
study.
Rhizophora mucronata
and
Avicennia maria
are
the dominant species in mangrove areas of the
Negombo estuary and the allometric relationship
between biomass and dbh (diameter at breast height)
of these species has already been determined
(Amarsinghe
and
Balasubramaniam,
1992b).
Nevertheless, investigations of TOC on mangroves in
Sri Lanka are non-existent.
1.3 Aims and objectives of the study
Present study is the first of this kind conducted on Sri
Lankan mangrove species with the objectives of
determining the pattern of total organic carbon (TOC)
distribution within the plant components, including
below ground components, through allometric
relationships between organic carbon in biomass of
plant components (stem, roots, leaves) and diameter at
breast height (dbh) of
B. gymnorrhiza
and
L.
racemosa.
Allometry has proven to be a useful method
not only to estimate total organic carbon sequestration
capacity of plant species, but also that of mangrove
ecosystems comprised of these species.
2 Results
2.1 Distribution of TOC among plant components
Except for the organic carbon in leaves of
L. racemosa
,
approximately half the biomass of all components in
the two species in the present study is composed of
organic carbon. Average organic carbon in woody
components (stems and branches) of
B. gymnorrhiza
was (10.17±4.52) kg/plant and it accounts for 53.5%
of total carbon available in the plant and this is lower
than that of
L. racemosa
(18.5±6.15 kg/plant) that
accounts for 71% of the total carbon in the whole
plant (Table 1). Although leaves of
L. racemosa
accumulate a relatively low amount of carbon
(0.79±0.2 kg/plant) and account only for 3.5% of the
total carbon in
B. gymnorrhiza
accumulates a
comparatively high amount of carbon in leaves (i.e.
(4.05±1.78) kg/plant and 21.3% of the total carbon in
the plant), revealing that
L. racemosa
accumulates
carbon predominantly in the sequestered form in
stems, branches and roots, and not as labile carbon in
the leaves (Table 1). Quarter of organic carbon in both
the species is accumulated in roots (under-ground and
below-ground together). The amount of organic
carbon retained in the above ground plant components
is about three times greater than that in below-ground
parts (Table 1).
Figure 1 presents the distribution of TOC among plant
components of trees with varying dbh. Although a
similar propensity in percentage TOC of stems,
branches and roots of
B. gymnorrhiza
observed with
increasing dbh, leaves showed a decrease with
increasing dbh values. Percentage TOC in branches of
L. racemosa
recorded a greater increase with
increasing dbh and decreasing values was recorded in
stem and roots with similar variations of dbh.
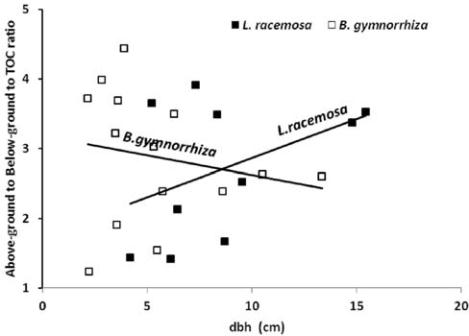
Relatively smaller trees (lower dbh) of
B. gymnorrhiza
accounted for a higher above (A) to below (B) ground
total organic carbon (TOC) ratio (A/B) and this was
observed to decrease with increasing dbh. On contrary,
L. racemosa
was revealed to account an A/B, that was
lower in smaller trees and it increases with tree dbh
(Figure 2).
Figure 2 Relationship between dbh and above-ground to below-
ground ratio (A/B)