Basic HTML Version



Rice Genomics and Genetics 2013, Vol. 4, No. 4, 14-27
http://rgg.biopublisher.ca
23
chemicals used for encapsulation. However, further
research is needed to optimize protocols for
production of androgenic viable synthetic seeds of
rice.
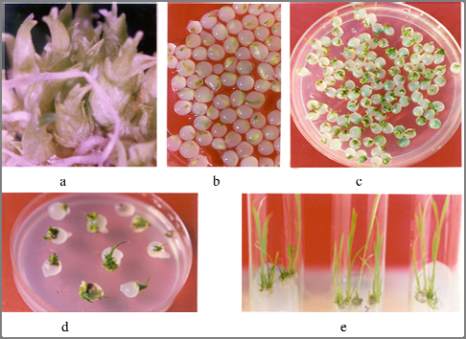
Figure 3 Pictorial depictions of synthetic seed production and
plantlet regeneration in rice (Roy, 2002)
Note: a: Stereomicroscopic view of germinating microtillers; b:
Encapsulated microtiller in sodium alginate (4%) beads; c:
Mass germination of beaded microtillers on MS medium with
no hormones; d: Seedling elongation from germinating
synthetic seeds; e: growing plantlets under in vitro culture
condition on MS basal medium
7 Limitations
Although results of intensive researches in the field of
synthetic seed technology seem promising for
propagation of crop plants. Several aspects of the
techniques are still underdeveloped and hinder its
commercial application:
1. High hand labour requirements and costly
procedures for the production of encapsulated
propagules.
2. Strain in recurrent production of somatic embryos
essential for synthetic seed production: Although large
quantities of somatic embryos can be rapidly produced
in many plant species, continuous supply is difficult as
the totipotency decrease with age of culture.
3. Improper maturation of the somatic embryos and
asynchronous development: Development of artificial
seeds requires sufficient control of somatic
embryogeny from the explants to embryo production,
embryos development and their maturation. Hence,
mature somatic embryos must be capable of
germinating out of the capsule or coating to form
vigorous normal plants. In some cases, somatic
embryos often develop extra cotyledons or poorly
developed apical meristems. This asynchronous
embryo development makes harvest difficult.
Uniformly mature somatic embryo development have
included physical separation of proembryonal cultures
to assure uniform callus size and physiological
synchronization by adding abscisic acid appears to
cause cell water (turgor) content to decrease, thereby
slowing embryo growth which inhibit germination of
embryos that would tend to germinate precociously.
4. Multiple somatic embryos development: Multiple
somatic embryos are often found on a single callus, in
which multiple stages of embryo development are
observed. This causes the non-uniform embryos to be
subjected to change nutrient conditions since the
nutrients are depleted by the developing tissues and
the replenished. Consequently many somatic embryos
have organs developing at different rates, which
contribute to asynchronous embryo development. In
some cases, this leads to precocious germination,
while in others the prevailing nutrient environment
may be conducive to shoot or root development but
not both.
5. Poor conversion of micropropagules into plantlets:
For commercial applications, somatic embryos must
germinate rapidly and should be able to develop into
plants at least at a rate and frequencies more or less
similar, if not superior to true seeds. To achieve
conversion of somatic embryos into plantlets and to
overcome deleterious effects of recurrent somatic
embryogenesis as well as anomalous development of
somatic embryos on their conversion, it is necessary to
provide optimum nutritive and environmental
conditions (Roy and Mandal, 2008).
6. Lack of dormancy and stress tolerance: Usually
somatic embryos continue to grow into seedling or
they revert back into disorganized callus tissue. This
inability to produce a resting phase where all embryos
are at the same arrested physiological and
morphological state also is a challenge to synthetic
seed development. Without this arrested growth stage,
synthetic seed cannot be successfully stored or treated